As of January 2004 au.riversinfo.org is archived under Precision Info
RIVERSINFO AUSTRALIA ARCHIVE
This archive provides non stylised versions of selected reference documents (formerly) resident on au.riversinfo.org.
A Guide to Diatoms as Indicators of Urban Stream Health
Dr Jacob John.
School of Environmental Biology Curtin University of Technology.
LWRRDC Occasional Paper 14/99 (Urban Sub Program, Report No.7) 2000
Contents
Foreword & Publication Details
Chapter 1
- Introduction
- How to collect, process and identify diatoms
- Phytoplankton
- Periphyton
- Permanent preparation
- Photomicrographs
- How to use this guide for identifying the diatoms
- Specialised literature for diatom identification
Chapter 2 Biology and Application of Diatoms
- Introduction
- Habit
- Habitat
- Frustule structure
- Cytology
- Cell division
- Movement of diatoms
- Life cycle
- Resting stages Ecology and application of diatoms Diatomite
- Diatoms in forensic science
- Classification and systematics
- Diatom research in Australia
Chapter 3 Features & Illustrated Identification
- Glossary for Diatom
- Glossary applicable to urban stream diatoms
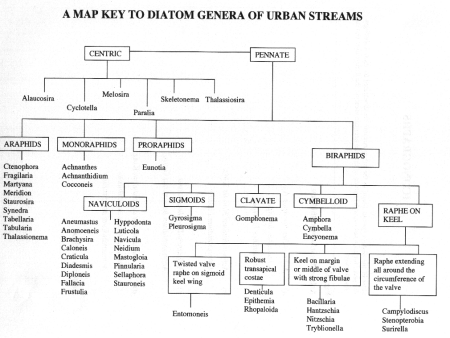
Chapter 4 Short Diagnostic Descriptions of Diatom Genera in Urban Streams
- I Centric Valves circular (radially symmetrical), no raphe many discoid or lobed plastids
- II Pennate diatoms Valves elongate, with or without raphe
Chapter 5 lconographs
References
Diatom Index
- Foreword
- Publication Information
Foreword
This report describes the outcomes of a research project conducted under the Urban Research and Development sub-program of the National River Health Program (NRHP).
The NRHP is an on-going national program established in 1993, managed by the Land and Water Resources Research and Development Corporation (LWRRDC) and Environment Australia. Its mission is to improve the management of Australia's rivers and floodplains for their long-term health and ecological sustainability, through the following goals:
- To monitor and assess the health of Australia's rivers.
- To enhance the management of river flows, water allocation and water use to ensure the sustainability of river and floodplain ecosystems.
- To encourage active management to improve the health of Australia's rivers, based on a sound understanding of the ecological and hydrological processes.
- To evaluate the effectiveness of river management actions at a national scale.
Urban streams and estuaries (i.e. those affected by runoff and discharges from urban areas) are an important subset of Australia's waterways. Most are degraded biologically, physically and chemically and therefore require appropriate methods to be developed for health assessment and management. The Urban R&D Sub-program, managed by the Water Services Association of Australia, comprises 8 research projects which were developed to meet research priorities for urban streams and estuaries within the goals of the NRHP and to complement existing NRHP projects on non-urban rivers. Thus, research focuses on development of standardised methods for assessing
the ecological health of urban streams and estuaries which can be linked with data on water and sediment quality. The urban R&D projects commenced in 1996.
The definition of health in urban waterways used is "the ability to support and maintain a balanced, integrative, adaptive community of organisms having a species composition, diversity and functional organisation as comparable as practicable to that of natural habitats of the region".
The eight projects of the Urban Sub-Program are:
| Decision support system for management of urban streams | Dr John Anderson
Southern Cross University, Lismore |
| RIVPACS (River InVertebrate Prediction and Classification System) for urban streams | Dr Peter Breen
CRC for Freshwater Ecology, Monash
University, Melbourne |
| DIPACS (Diatom Prediction and Classification System) for urban streams | Dr Jacob John
Curtin University, Perth |
| Sediment chemistry- macroinvertebrate fauna relationships in urban streams | Dr Nick O'Connor
Water EcoScience, Melbourne |
| Classification of estuaries | Dr Peter Saenger
Southern Cross University,Lismore |
| Literature review of ecological health assessment in estuaries | Mr David Deeley
Murdoch University, Perth |
Estuarine health assessment using benthic macrofauna
| Dr Gary Poore
Museum of Victoria, Melbourne |
| Estuarine eutrophication models | Dr John Parslow
CSIRO Marine Laboratories, Hobart |
Publication Details
Published by:
Land and Water Resources Research and Development Corporation
GPO Box 2182 Canberra ACT 2601
Telephone: (02) 6257 3379
Facsimile: (02) 6257 3420
Email: public@Iwwrrdc.gov.au
WebSite: www.lwrrdc.gov.au
© LWRRDC
Published Electronically on au.riversinfo.org by the Environmental
Information Association (Incorporated) with the permission of LWRRDC and
Environment Australia. Environment Australia assisted by providing copies
of the manuscript for electronic publication. The Natural Heritage Trust
provided project funds which were used to assist in publishing this material. In the case of variation between this document and the hard copy original the original takes precedence. (Bryan Hall).
Disclaimer:
The information contained in this publication has been published by
LWRRDC to assist public knowledge and discussion and to help improve the
sustainable management of land, water and vegetation. Where technical information
has been prepared by or contributed by authors external to the Corporation,
readers should contact the author(s), and conduct their own enquiries,
before making use of that information.
Publication data:
A Guide to Diatoms as Indicators of Urban Stream Health. Dr Jacob John. School of Environmental Biology Curtin University of Technology.
LWRRDC Occasional Paper 14/99 (Urban Sub Program, Report No.7) 2000
ISSN 1320-0992
ISBN 0 642 26764 2
Managing Agencies
-
The Urban Sub Program was managed by Water Services Association of Australia
under a contract with Land and Water Resources Research and Development
Corporation.
-
Environment Australia funded the research projects of the Urban Sub Program
of the National River Health Program.
-
Hard copy designed by: Nuttshell Graphics, North Melbourne
-
Hard copy printed by: Aristoc Offset, Mt Waverley
- Electronically Published by: Environmental Information Association (Incorporated)
Acknowledgements
I wish to express my gratitude to Dr John Langford (Executive Director, Water Services Association of Australia) for his constant encouragement in completing the project.
My thanks are extended to Water and Rivers Commission, WA for financial support for the water chemistry involved in this project.
I wish to thank the School of Environmental Biology, Curtin University for providing facilities for the research project and Miss Fiona Butson and Miss Erin Lowe, my PhD students and Ms Jean Burgess of the School of Environmental Biology for their assistance in preparing this guide.
I acknowledge the assistance of research assistant Mr Chris John Siva for his general assistance in the project.
Chapter 1 Introduction
- Introduction
- How to collect, process and identify diatoms
- Phytoplankton
- Periphyton
- Permanent preparation
- Photomicrographs
- How to use this guide for identifying the diatoms
- Specialised literature for diatom identification
The ultimate 'monitor' of aquatic systems such as urban streams is 'aquatic life' itself. No other single taxonomic group of aquatic organisms is as sensitive as diatoms to changes both in water quality and habitats. For any biomonitoring program to be successful, the biomonitors need to be identified accurately. Lack of practical guides to identify diatoms is generally acknowledged as a problem limiting their use as biomonitors in Australia. This publication is an attempt to address this problem.
Only very few diatoms can be identified to the species level by examining live cells. With experience, most of the genera can be identified on the basis of shape of cells, habit (single, colonial), habitats and chloroplastids. The cells have to be cleared of organic materials and converted into permanent slides for accurate identification to species level. The slide collection becomes an invaluable resource for assessment of stream health.
Periodical collections from the sites provide a series of snapshots of stream health – very much like a family photo-album, not for an occasional nostalgic look, but for appropriate action to rectify the maladies diagnosed by the assessment and to see whether any remediation measures undertaken are really working.
How to collect, process and identify diatoms
Collection techniques and preparation of permanent slides are described in detail in the volume 'Diatom prediction and classification system for urban streams' (National River Health Program; Urban sub program 2000). Here I wish to make only brief comments on collection, permanent preparation techniques and general guidance on the use of the diatom atlas presented here so that this publication can be used independently.
Phytoplankton
Although fast flowing streams have very few plankton, stagnant pools and slow flowing streams, abundant in Australian urban areas may be rich in phytoplankton. Diatom blooms are common in these streams. A phytoplankton net (25µm-mesh size) is ideal for collecting such samples. Quantitative sampling requires collection of a specific volume of water and subsequent concentration and enumeration.
Periphyton
The attached microalgae of which diatoms are a dominant component, can be collected by scraping natural substrates in streams. Stones (pebbles) and macrophytes are good sources of diatoms. Floating filamentous algae or floating wood and plant materials are often full of loosely attached diatoms. Diatoms grow attached to concrete substrates in urban drains. Epiphytic diatoms grow attached to macrophytes and large algae. Metaphyton includes diatoms loosely attached to algal mats and filamentous algae. Typically diatoms can be seen in the form of brown or black, slimy coatings on stones, submerged plants and mud. The coloured coatings may be removed with a brush, 'spoon' or blunt scalpel from hard substrates and by squeezing or washing off from soft substrates like macrophytes and filamentous algae.
Benthic diatoms may be collected by 'scooping' the top 3–5 cm of the sediment of a stream, with a vial but this method disturbs and mixes the sediments. An intact 'core' may be collected, by pushing an inverted open vial (with a small hole at the bottom), into the sediment and lifting it up. Using this quantitative method you may collect epipelon (growing on mud) and episammon (growing on sand) intact from a specific area. Motile species are common in mud samples.
Quantitative sampling requires a constant surface area of substrates for collection to minimise variation due to sampling errors. Artificial substrates (e.g. JJ Periphytometer) used in the current project collect a representative samples of epiphytic (growing on macrophytes), epilithic (growing on stones) and epipelic (mud) diatoms, amenable to quantitative analyses (John 1998; 2000).
Permanent preparation
Fresh samples should be examined, if habit and chloroplasts are to be observed. 'Colonies' are disintegrated by the processing employed for permanent preparation. Chloroplasts are destroyed by the chemical processing required for making permanent slides. If diatom samples are preserved by Lugol's solution, 4% formalin or Transeau's Algal Preservative (6:3:1 = Water, Ethyl alcohol, Formalin), colonial structure and chloroplasts can be retained.
Concentrated diatom samples are boiled in 50 to 60% Nitric Acid to oxidise the organic materials and then the acid is washed off with deionised water by settling or centrifugation. The final 'cleared' diatom samples – an ash coloured material is mounted in a mounting medium e.g. Napthrax with a refractive index close to 1.7 and placed on a hotplate until the solvent in the medium evaporates.. The slide is cooled immediately, by removing from the hotplate. The diatom slide thus obtained is permanent (see details in Chapter 3: Diatom Prediction and Classification System for Urban Streams 2000).
Photomicrographs
The cleaned cells in permanent slides are observed under 40X or oil immersion (100X) and photographed using black and white Tech.Pan Kodak® film for best results. All the micrographs in this publication have been produced this way.
How to use this guide for identifying the diatoms to species level
An appreciation of the biology and application of diatoms is highly recommended for anyone contemplating using diatoms as biomonitors, irrespective of his or her background (biologist, geographer, botanist, zoologist, engineer or environmental scientist).
The second chapter is a general introduction to the biology, ecology and application of diatoms from an Australian perspective. Marine and freshwater diatoms are treated together. The third chapter gives you a taste of the basic vocabulary one needs to build up as well as some helpful hints on features relevant to diatom identification. Chapter four is a brief diagnostic description of 53 genera treated in this publication: each illustrated with one or more micrographs. The chapter concludes with a summary key or 'mind map' of the genera. I have not attempted any dichotomous keys because I believe conventional dichotomous keys for diatom identification are time consuming and not user friendly. A quick reference to the 'map' provided at the end of Chapter four is easier to use than a conventional key. You should be able to identify most of the diatoms in temperate urban Australian streams to generic level, using the illustrated key. Once you identify the diatom to generic level, you may go directly to the iconographs of the species which are arranged according to the key to genera in alphabetical order and examine the micrographs of the species of the genus in question. Hopefully you will be able to identify a diatom. In case the species you want to identify is not in this publication (it is likely that there will be some), you may refer to the literature mentioned below, specifically John 1983a and 1998, or you may contact me by mail or email and forward the specimen or a good photograph. As the honorary curator of Curtin University Diatom Herbarium, I will try and identify the specimen for you. Curtin University International Diatom Herbarium houses the largest collections of diatoms in Australia. I certainly welcome any additions to our collections
Specialised literature for diatom identification
Archibald (1983); Bateman & Rushforth (1982); Camburn, Kingston & Charles (1984-86); Cassie (1989); Clark & Rushforth (1977); Czarnecki & Blinn (1978); Dodd (1987); Domitrovic & Maidana (1997); Foged (1976, 1978, 1984, 1987);Frenguelli (1923- 1924 reprint 1976); Gasse (1986); Germain (1981); Hansman (1973); Hustedt (1930, 1927-1966);John & Economou-Amilli (1991); John & Van Riessen (1992); Krammer & Lange Bertalot (1986, 1988, 1991);Lange-Bertalot & Krammer (1989); Lange-Bertalot & Metzeltin (1996); Lange-Bertalot (1979, 1996); Lange-Bertalot et al. (1996); Metzeltin & Lange-Bertalot (1998); Metzeltin & Wilkowski (1996); Patrick & Reimer (1966, 1975); Podozorski & Hakansson (1987); Podzorski (1983); Reichardt (1984); Round, Crawford & Mann (1990); Schmidt et al. (1874-1959); Schoeman & Archibald (1976 – 1979); Schoeman (1973); Sims (1996); Snoeijis & Vilbaste (1994); Snoeijis (1993); Wilkowski (1994); Williams & Round (1987)
The following references are on Australian diatom flora.
Foged (1978); Thomas (1983); Hodgson, Vyerman & Tyler (1997); Holland & Clark (1989); John (1980, 1981a, 1981b, 1981c, 1982a 1982b, 1983a, 1983b, 1986, 1987 1988, 1989, 1990a, 1990b, 1990c, 1990d, 1991, 1992, 1993a, 1993b, 1993c, 1994, 1998); Vyerman et al. (1995)
Chapter 2 Biology and Application of Diatoms
- Introduction
- Habit
- Habitat
- Frustule structure
- Cytology
- Cell division
- Movement of diatoms
- Life cycle
- Resting stages Ecology and application of diatoms Diatomite
- Diatoms in forensic science
- Classification and systematics
- Diatom research in Australia
Introduction
Diatoms are unicellular microalgae universally distributed in all types of aquatic environment. The most obvious feature of diatoms is their siliceous cell wall (frustule) made of two intricately sculptured halves (valves). Diatoms form the majority of floating algae (planktonic) in marine and fresh waters. It is estimated that these microalgae may account for 40 to 45% of oceanic production making them more productive than all the world's rainforests (Mann 1999). It is not surprising that they play an important role in global cycling of Silica and Carbon and at the same time sustain fisheries (Mann 1999). Diatoms multiply rapidly, maintaining a dynamic population of varying size. They are diploid ( double set of chromosomes) and show high variation in sexual reproduction and life cycle patterns.
Diatoms are the most species rich group of algae with tens of thousands of species (Mann 1999). There are currently over 260 genera of living diatoms with over 100 000 species (Round et al. 1990). Diatom taxonomy is in a state of revolutionary change involving splitting of genera, creation of new ones and revision of criteria for classification. With molecular genetics and breeding as added tools, taxonomy of diatoms has become a dynamic field of research. This has added to the value of diatoms as biomonitoring tools of aquatic systems, which has been recognised world wide (John 1993b, 1998). Diatoms have been widely applied in palaeoecological reconstruction of past environmental conditions. Accurate identification of taxa with information on geographical distribution is essential for the use of diatoms as reliable indicators.
Diatoms are distributed throughout Australia in marine, freshwater, saline and soil environments in abundance. In keeping with a worldwide trend, there has been a steady increase in diatom research in Australia over the past twenty years. This chapter aims at providing an overview of the biology and applications of diatoms.
The main distinguishing features of diatoms
- Unicellular or cells connected together to form chains, each cell is protected by a pair of siliceous cell walls (frustule) covered by organic material. The cell walls form a box with two equal halves called valves (Fig. 1). The valves are connected by a 'cingulum' (girdle) made of thin siliceous girdle bands termed copulae.
- No flagellate cells except spermatozoids in the centric diatoms
- The chloroplasts in diatoms are usually golden-brown: with pigments: Chl a and c2, (1-3 Carotene dominated by fucoxanthin – neofucoxanthin, diadinoxanthin and diatoxanthin. The chloroplast DNA is organised into a ring shaped nucleoid.
- The life cycle involves, vegetative cell division and sexual reproduction by isogamy (fusion of similar gametes), anisogamy (fusion of dissimilar gametes) or oogamy (fertilization of egg by spermatozoid).
- Stored food materials: chrysolaminarin and oil
Habit
Functionally diatoms are unicellular but several taxa form chains or filaments due to daughter cells remaining connected even after cell division, often by polysaccharide pads or linking spines (Fig. 2a & b). The colonies may be straight, zig zag, fan shaped, star shaped, or radiate. Then, there are motile colonies of Bacillaria paxillifer with a characteristic 'slide rule' like movement. Dichotomously branched mucilaginous stalks are characteristic of few species (Fig. 3c). There are tube dwelling colonial diatoms, the tubes assuming characteristic tree-like branching. The marine epiphytic species Mastogloia cocconeiformis associated with stromatolites in Shark Bay in Western Australia secrete individual tubes for each frustule, the daughter cells initiating lateral branches of tubes after each cell division (Fig.3 a, b & d). The tubes are made of mucopolysaccharides and form a highly ramifying network (John 1993a).
| Fig. 1 Frustule structure and symmetry of diatoms |
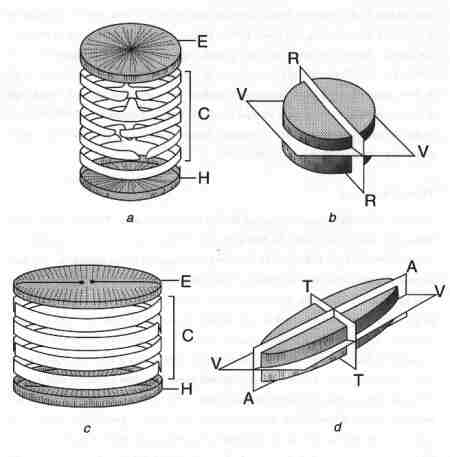
|
- Centric diatom: E = epivalve, C = cingulum, H = hypovalve. The cingulum (girdle) consists of copulae (girdle bands). The copula adjacent to the valve is valvocopula.
- Centric diatom symmetry: VV = Valvar plane, RR = Radial plane.
- Pennate diatom: E = epivalve, C = cingulum with cingular elements (girdle bands), H = hypovalve. Valve face is the flat surface of the valve up to the margin; Mantle is the steep edge (side) of the valve.
- Pennate diatom symmetry: VV = valvar plane, AA = apical plane, TT = transapical plane. Apical axis connects the two poles through the median line. Transapical axis connects the centre of the epivalve to that of the hypovalve.
|
Habitat
Diatoms occupy a wide variety of habitats from floating (planktonic) to attached (periphyton). The latter may be attached to rocks (epilithon), sand (episammon) mud (epipelon) or plants (epiphytic) (Fig. 2f). The term metaphyton is used for diatoms, which are loosely attached to macrophytes or other submerged substrates. One of the most rare habitats of the diatoms is inside the empty tubes of setae of dead Artemia colonised by a species of Navicula (Fig. 2c) (John 1994). Further adding to the diversity of habitat is the epizoic diatom (growing on living animals reported in several zooplankton) (John 1991) (Fig. 2d). There have also been reports of diatoms growing on the feathers of birds. A few members of benthic foramnifera harbour endosymbiotic diatoms which lack siliceous frustules or have only vestigal frustules (Lee & Xenophontes 1989).Mucilage ensheathment secreted by the charophyte Nitella hyalina is colonised by special assemblages of diatoms (Fig. 2e) (John 1994).
Frustule Structure
A diatom cell under the microscope may be seen in different views. The term 'valve view' refers to the view of the 'face' of a diatom frustule. The side view (profile) is referred to as the girdle view (Figs. 1 & 4). The symmetry and structure of the frustule is extremely important in the systematics of diatoms. The symmetry is basically related to their axes. The apical axis connects the two poles of valve, and the transverse axis lies perpendicular to the apical axis. The transapical axis connects the central point of one valve to that of the other. Three planes can be recognised in relation to the three axes. The valvar plane is parallel to the valve face and cell division takes place in this plane. The apical plane runs through the apical axis and the transapical plane runs through the transapical axis. Although diatoms exist in different shapes, broadly there are two main groups: centric and pennate. Most of the centric diatom frustules are radially symmetrical, whereas the majority of the pennate diatoms are bilaterally symmetrical with many exceptions. Genera with complex shapes include Entomoneis, Surirella, Campylodiscus, Amphora, Epithemia and Rhopalodia (Fig. 4). The majority of the pennate diatoms have a longitudinal slit termed raphe (Figs 1, 4 & 5). The pennate diatoms with raphe are referred to as 'raphids' and without raphe as 'araphids'.
The upper lid of the frustule (epivalve) overlaps the lower lid (hypovalve). The epivalve and hypovalve are connected by girdle bands termed copulae (Figs. 1 & 4) made of silica. The copulae associated with the epivalve is termed epicingulum. Hypocingulum is the term applied for the bands associated with the hypovalve. The bands not only hold the valves together to protect the cell, but are also involved in accommodating the cell volume increase during the cell cycle. Three types of girdle bands are encountered in diatoms. (1) scales (2) split rings and (3) closed rings (Stosch 1975). Scales are considered to be more primitive than the rings which are more common in the vast majority of diatoms. The closed bands are considered the most advanced. The copula adjacent to the valve ('valvocopula') is underlapped by the valve edge. The structure and type of bands are significant in the taxonomy of diatoms. In some species of diatoms, the copulae have inwardly directed septa (Fig. 4 f & g). In the genus Mastogloia the valvocopula has internal chambers termed partecta (Fig. 4 i & j).
The siliceous structure of the valve consists of transverse and short lateral ribs separated by tiny 'pores' or 'areolae' (puncta) (Figs. 5 & 6). These little holes arranged in rows are termed stria (striae). Secondary deposition of silica may transform the areolae into complex 'chambers' or locules as seen in centric diatoms. The locules are occluded externally or internally by siliceous flaps or porous plates termed vela. 'Vela' may be classified into different types. In a few raphid diatoms the pores may be occluded by a very delicate layer of silica termed 'hymen' with tiny elongate perforations.
In addition to the simple and complex pores on the valve, clearly demarcated finer pores in groups, surrounded by a clear rim are observed in the poles of a few species. The term 'ocellus' is applied to this cluster of fine pores e.g. Auliscus (Fig. 4 k). Many araphid and raphid diatoms do have small pores at their poles, termed pore field; These pores are involved in the secretion of mucilage (Fig. 5 d & e). Diatom valves may have spines, hair-like structures (setae) and horns which may be solid or hollow. These siliceous appendages from the frustules provide linkage between cells to form chains (Fig. 2a). The hollow setae of the marine genus Chaetoceros may have cytoplasm and even chloroplastids. In addition to these, many species have special processes termed 'portules' (processes) (Fig. 6). There are two types of portules: fultoportulae (strutted processes) and rimoportulae (labiate processes), the former is confined to the centric order Thalassiosirales and the latter wide spread in centric and pennate diatoms. Scanning Electron Microscopy (SEM) is required to observe these structural details. However light microscopy is sufficient to identify the common species of centric diatoms in urban streams.
| Fig. 2 Diatom habits and habitats |
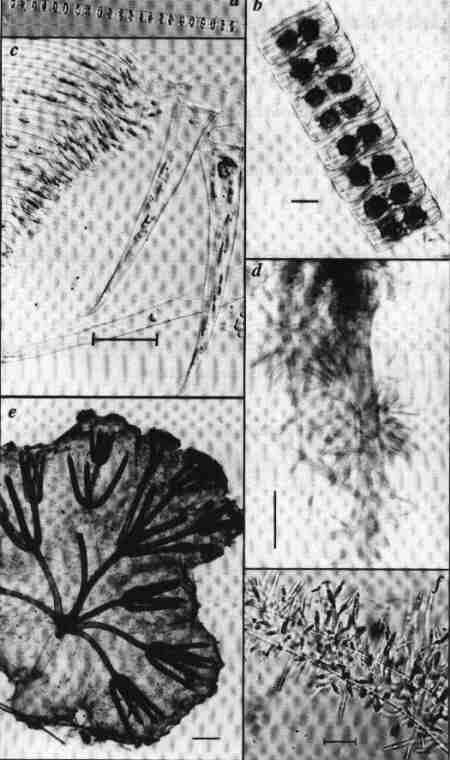
(larger image)
|
|
Scale bar = 10 µm
- A chain forming planktonic diatom (Skeletonema costatum), the cells are linked by siliceous appendages originating from the margin of the valves.
- An epiphytic colony of diatoms (Achnanthes brevipes); the cells are connected by the valve face.
- A Navicula sp. colonising the setae of dead Artemia in a salt lake.
- An epizoic diatom (Falcula hyalina) growing on the appendages of living copepods from the Swan River estuary, Western Australia.
- The mucilage ensheathment around the growing shoots of Nitella hyalina – a charophyte colonised by an assemblage of diatoms.
- An epiphytic diatom with mucilaginous stalk (Brachysira aponina) growing on the tubes secreted by another species of diatom (Mastogloia cocconeiformis) from Shark Bay, Western Australia.
|
| Fig. 3 Habits and habitats of diatoms |
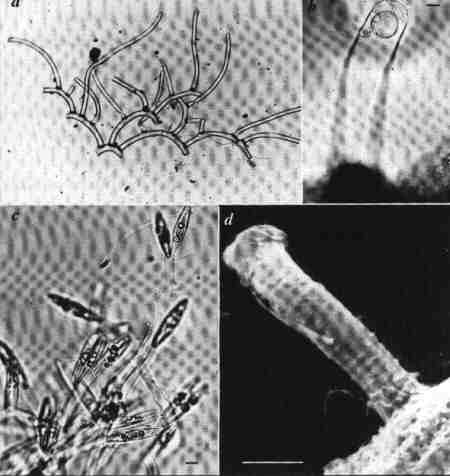
(larger image)
|
- The tube dwelling diatom Mastogloia cocconeiformis; each branch is secreted by a newly divided daughter cell.
- The tube dwelling diatom inside the mucilaginous tube. Scale bar = 10 µm
- The diatom Brachysira aponina with dichtomously branched mucilage stalks. Scale bar = 10 µm.
- A Scanning Electron Micrograph (SEM) of the mucilaginous tube of Mastogloia cocconeiformis. Scale bar = 10 µm.
Both species above are associated with stromatolites at Shark Bay, Western Australia.
|
| Fig. 4 Diatoms in varying shapes and symmetry |
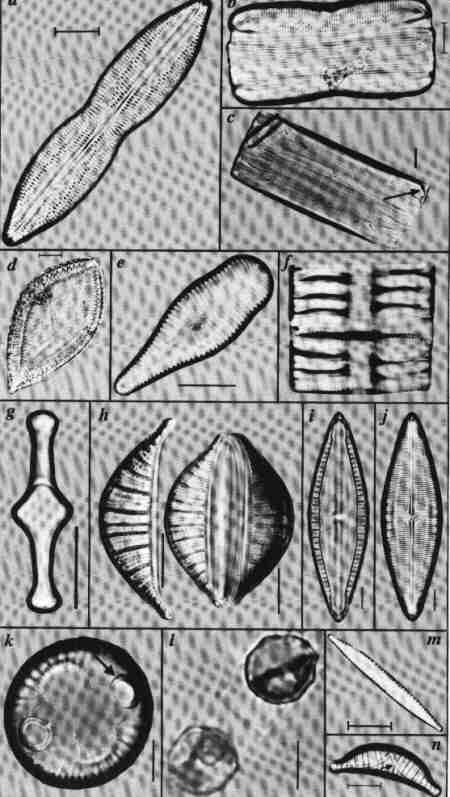
(larger image)
|
|
Scale bar = 10 µm
- Valve view of a bilaterally symmetrical diatom (Parlibellus panduriformis), showing the raphe and striae; both poles are similar (Isopolar).
- The girdle view of the above; note the valve mantle and the cingulum
- The girdle view of Eunotia (a genus with a short reduced raphe visible only in girdle view at the poles)
- A heteropolar diatom (differs in morphology of poles) Surirella; The raphe is located on a raised keel on the whole margin (circumference).
- A clavate (club shaped), heteropolar diatom (Gomphonema)
- Girdle view of a diatom (Tabellaria) showing septa (flat extensions of girdle bands)
- The valvocopula of the above species showing the septum in valve view
- Bilaterally assymmetrical, lunate (crescent shaped) dorsiventral valve of Rhopalodia; the raphe is located on the dorsal margin on a keel; the valve surface is crossed by robust costae and the frustule of Rhopalodia with the two valves.
- A valve of Mastogloia showing the teeth-like locules (Partecta) of its valvocopula
- The same valve with the striae on focus
- The marine diatom Aliscus showing the ocellus – a cluster of closely arranged pores.
- Resting spores of a marine planktonic diatom (Chaetoceros)
- A diatom with marginal raphe on a keel supported by Fibulae.
- An dorsiventral, arcuate shaped diatom with the raphe on an arched keel (Epithemia).
|
| Fig. 5 Structure of Pennate diatom frustule. Scanning Electron Microscopic (SEM) views. |
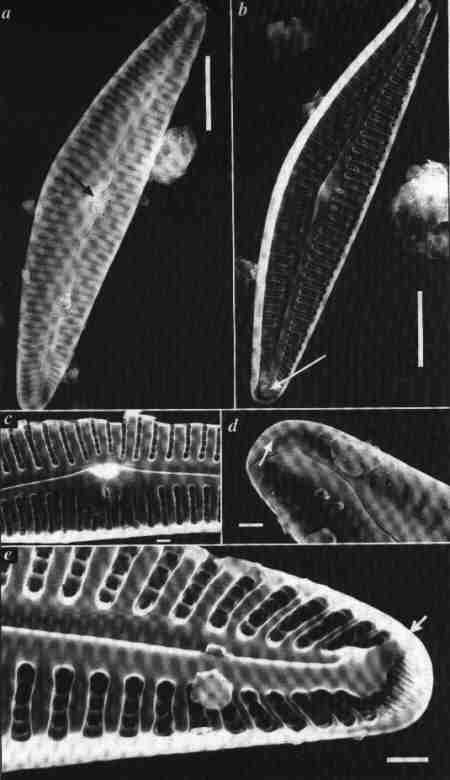
(larger image)
|
- External surface of a bilaterally asymmetrical, isopolar, dorsiventral biraphid diatom (Cymbella). Arrow points to central nodule; lateral to the nodule on the ventral side, 4 isolated pores (stigmata) are seen; note the dorsal and ventral uniseriate striae (made of single row of areolae) and central pores of raphe. Scale = 10 µm
- Internal surface view of the valve showing uniseriate striae, note the structure of areolae, the internal opening of the stigmata, the raphe-sternum (siliceous axial area in which the raphe is embedded) and the polar nodules. Scale = 10 µm
- Internal surface view of the centre: The central endings of the raphe are hidden. Scale = 1 µm
- External surface of a pole showing the raphe ending (terminal fissure) curved towards the dorsal side and clusters of small polar pores. Scale = 1 µm
- Internal surface view of a pole showing the polar ending of the sternum and the small pores. Scale = 10 µm
|
| Fig 6 Structure of a centric diatom frustule (Thalassiosira australis) |
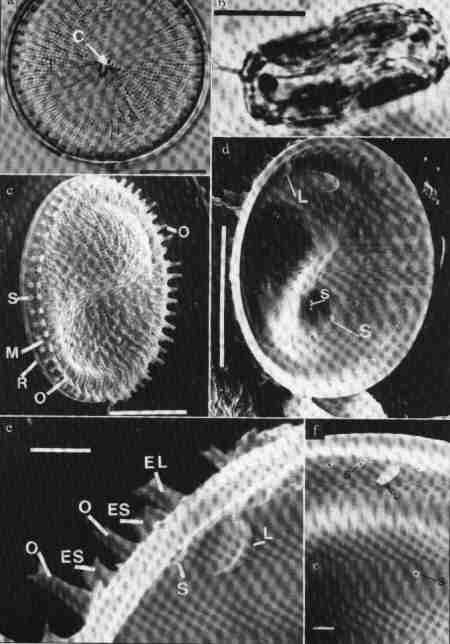
(larger image)
|
- Valve view (Light Microscopy) showing the radiate arrangement of areolae and the undulate valve face.
- Girdle view of frustule drum-shaped, undulate (Light Microscopy) Scale = 10 µm
- SEM structure of valve, External surface view
O = Occluded process; R = Rim of mantle; S = Strutted process; M = Mantle. Scale = 10 µm
- Internal surface view of valve
S = Strutted process L = Labiate process Scale = 10 µm
- SEM view of the mantle, O = Occluded process
L = Labiate process (internal surface view), EL = External labiate process; S = Strutted process, ES = External strutted process
- Enlarged view of internal surface
S = Strutted process; L = Labiate process Scale = 1 µm
|
| Fig. 7 Plastids of diatoms |
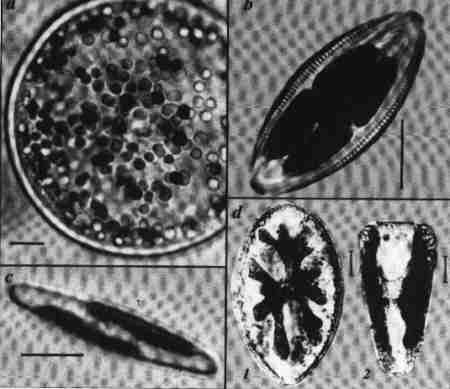
(larger image)
|
- Scale = 10 µm
- A centric diatom (Hyalodiscus), with many disc-shaped chloroplastids.
- An H shaped plastid connected by a central bridge in Amphora.
- Two staggered plastids each appressed against girdle. Navicula.
- Surirella.
- Highly lobed large chloroplastid under each valve connected by a central link. Valve view.
- Girdle view showing the location of the plastids.
|
| Fig. 8 Cell Division |
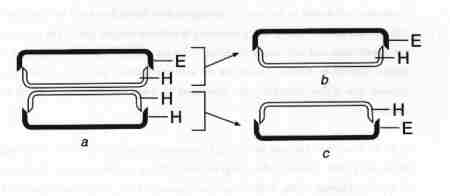
(larger image)
|
- A diatom cell dividing; only valves are shown. E = Epivalve, H = Hypovalve.
- Products of cell division; a daughter cell maintaining the same size as the parent, with a newly formed hypovalve.
- The second daughter cell with the parent's hypotheca acting as its epitheca, with a new hypotheca. This daughter cell is smaller than the other.
|
(larger image)
|
- A valve view of a naviculoid diatom with an 'H' shaped (Lyrate) hyaline area connected by a central nodule (arrow) SEM.
- Valve view of a diatom with short raphe (not seen in valve view) and a dorsiventral outline with undulations on the dorsal side (Eunotia) (Light Microscopy).
|
Cytology
Unicellular and diploid as diatoms are, cytologists have found them ideal for studying cell structure and mitosis. However, the siliceous cell wall has been the main focus of attention for classification and systematics of diatoms. Living diatoms have to be cleared of the organic material by boiling in acids and the siliceous frustules have to be mounted in a medium with higher refractive index than glass or water, for observation of cell wall details.
In recent years, there has been an increased awareness of the need to describe the structure of the living cells of diatoms, the protoplast, especially the plastids as part of the diagnosis of species. Plastids are the most conspicuous feature of living diatoms and their number and shape are consistent features of taxonomic importance. Cox (1996) in an attempt to formulate a key for identification of living diatoms, has described in detail the nature of chloroplastids in several groups of diatoms.
Chloroplastids may be rounded or lobed discs or large plate-like with or without lobed margins (Fig. 7), and may range from one to two, four or more. A typical centric diatom has many disc shaped plastids, arranged close to the periphery surrounding a large central vacuole or scattered throughout the cell. The raphid diatoms tend to have large chloroplasts (one to four), lying along the girdle with central nucleus flanked by two large vacuoles. Pyrenoids (starch bodies) one or two or many, may be found per chloroplast.
Diatoms store Chrysolaminarin, b 1-3 glucan and lipids as their food reserve. Lipids may be stored in high quantity, with a high percentage of polyunsaturated fats. Feeding fish and crustaceans with diatoms with high quantities of unsaturated fatty acids has commercial advantages and there is considerable interest in culturing such diatom species for aquaculture industry (Lokman 1998).
Cell Division
Vegetative cell division is the most common method of reproduction in diatoms. Like the cell cycle of any other eukaryote, diatoms must synthesise double the number of mitochondria, plastids, dictyosome and other organelles, and replicate their chromosomes before cell division. The full set of cell components should be apportioned equally to the daughter cells and then the daughter cells should make new cell walls with the same pattern of areolae as the mother cell. However the cell walls of the mother cell are not discarded, instead, they are utilised by the daughter cells. The rigid structure of the cell walls impose certain restrictions on the growth of the daughter cells. The epitheca (the overlapping cell wall) of the mother cell is retained by one of the daughter cells which synthesises a new hypotheca. The hypotheca of the mother cell then functions as the epitheca of the other daughter cell which in turn makes its own hypotheca. The two thecae of the parent cell thus become the epithecae of the daughter cells. The result of this unique cell division is that one of the daughter cells becomes smaller than the other (Fig. 8). As cell division continues over a period, a proportion of the population continues to get slightly smaller. However not all species of diatoms experience the diminution in size through cell division; a few have fairly elastic girdle bands which allow growth of the cell walls similar to the parents' cell walls.
The cell cycle of diatoms may last 8 to 24 hours in most cases, depending upon the species and environmental conditions. For example Thalassiosira species may divide rapidly taking only 8 hours. Pickett-Heaps, the Australian cytologist and colleagues have contributed substantially to our understanding of mitosis in diatoms (Pickett-Heaps et al. 1980; Pickett-Heaps 1987).
Movement of Diatoms
Pennate diatoms with raphe display a characteristic jerky movement forward or backward, different species taking different path patterns. The path patterns may be curved, circular, straight or zig zag, leaving trails on the substrates. The raphe should be in contact with the substrate for movement to occur. Although the general velocity of the diatom movement is less than 1 to 25µm/s, they can accelerate at times to 100-200 µm/s and come to a sudden stop. The involvement of raphe in movement has been recognised for a long time. The trails left by diatoms have been the subject of research for several years. The position of the raphe in the diatom varies: the majority have their raphes in the median apical plane, a few have laterally placed raphes on a keel in the valve margins and still others have raphe only in one valve (Figs. 1, 4, 5, 8 d, e, 9 a & b). However all raphes open to both external and internal faces of the valve, and form channels of interactions with the substrates. Secretion of mucilage through the narrow channels of the raphe from the cell to the substrates form the basis of modern theories of diatom movement. Ultrastructural studies of diatom cells, specifically raphe, cytoskeleton, plasmalemma and vesicles have provided the structural basis for the modern theories of locomotion. Cytoplasmic streaming and jet propulsion once proposed as driving the motion, are only of historical interest. The theory based on polysaccharide fibres extruded through the raphe and driven by the interaction of microfilaments and the plasmalemma provides the most reasonable explanation for diatom locomotions matching with structural details of diatom cells (Edgar and Pickett-Heaps 1983; Round et al. 1990).
Life cycle
The life cycle of diatoms involves cell division, formation of resting spores, asexual vegetative enlargement and sexual union followed by auxospore formation. While cell division as a method of reproduction is common to all diatoms, the other stages mentioned above may not be universally applicable to all diatoms and may occur only under certain circumstances. Information on aspects of sexual reproduction is confined to 200 species out of 20,000 species of diatoms currently known. Only a small proportion of diatom populations in a given environment is known to undergo sexual reproduction. Sexual cycle may involve in some instances up to 40 years (Mann 1999) and annual sexual cycle is known only in few. Sexuality in diatoms is the means of size restoration and genetic recombination rather than dormancy or multiplication. Resting spores obviously are produced as a means of dormancy (Edlund & Stoermer 1997).
The cells must have a minimum size range about 40% of its maximum size before they can sexually reproduce. Conditions such as the stage of cell cycle, temperature, lights, nutrients, hormones, trace metals and osmolarity should be suitable for initiation of sexual reproduction. The diminution in size caused by series of vegetative mitotic division is rectified by auxospore formation following sexual exchange of genes. Sexual exchange in centric diatoms is by oogamy and is relatively uniform compared to pennate diatoms.
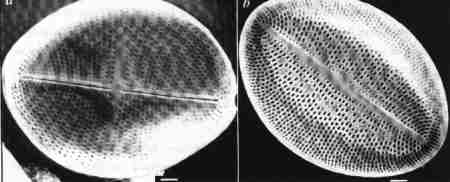
Oogamy in centric diatoms involves the fertilisation of one or two non-motile oogonia by motile uniflagellate spermatozoids. The zygote then develops into an auxospore from which large sized cells are formed (Fig. 9 c, d & e). On the other hand, pennate diatoms sexually reproduce by isogamy (fusion between identical gametes). Physiological or behavioural anisogamy is observed in a few pennate diatoms (Edlund & Stoermer 1997; Mann 1999). Rarely, gametes produced by the same vegetative cell unite to form a zygote (Paedogamy). Other variations reported include autogamy (fusion of haploid nuclei after meiosis before cleavage of the gametangium) and parthenogenesis (Round et al. 1990; Mann 1999).
Resting Stages
Resting stages ensure survival of species of diatoms only when conditions are suitable. Resting stages include morphologically distinct spores and physiologically distinct cells. Limiting nutrients such as nitrogen and phosphorous, low light and temperature may induce resting spore formation of planktonic diatoms such as Chaetoceros in marine waters (Fig. 4 l). The spores are denser than vegetative cells and sink to the bottom of the water when a bloom of Chaetoceros terminates. The resting cells and spores are often considered overwintering forms but are able to provide 'seed stocks' for subsequent blooms. Top sediments (2cm) of any estuary or marine body or stagnant waterbody would have spores and resting cells of many diatoms remaining viable for many years. The resting cells germinate from the sediment and seed the standing water under suitable environmental conditions. Seasonal succession of planktonic diatoms is linked to the resting stage.
The resting stages are also common in terrestrial and aerophilous diatoms. They remain dormant to survive desiccation, but are viable for a long period. Often the resting cells are morphologically similar to the vegetative cells but have very thick cell walls and may have various spines and warts (Round et al. 1990).
Ecology and application of diatoms
Large number of species (between 104 and 105) with distinct range of habitat and tolerance to environmental conditions and the siliceous nature of the cell wall make diatoms the ideal ecological indicators with a wide range of applications both as living organisms and fossils.
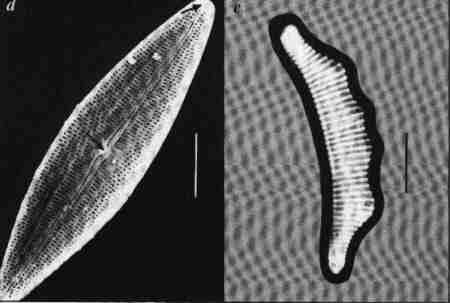
| Fig. 9 |
(larger image)
|
- Cocconeis – a monoraphid diatom. Raphe Valve (RV); note the sample, straight raphe with terminal pores.
- Cocconeis – Raphe Less Valve (RLV) note the axial area.
|

(larger image)
|
- Auxospore formation in the centric diatom Melosira.
- & e. Straight chain of cells in pairs formed from the auxospore (Melosira) note all the daughter cells from the auxospore are large.
|
Diatoms have been used as indicators of environmental change in flowing waters and lakes. Periphytic diatoms of lakes respond to eutrophication. Quantitative data on the ecological optima and tolerance of diatoms along a gradient of lake trophic status are generated from data sets of surface sediment diatom assemblages. Multivariate analyses are employed to determine the environmental factors most strongly correlated with species distribution. Several studies have demonstrated that the distribution of diatom taxa in surface sediments is correlated with lake trophic status (Fritz et al. 1993; Hall & Smol 1999).
After determination of the dominant environmental factors influencing diatom distribution, transfer functions can be generated to infer environmental variables from diatom assemblages. Past trophic conditions of a lake can be determined from fossil diatom assemblages. Such diatom inference models have been developed for estimating total N in Ontario lakes, Canada (Christie & Smol 1993). Historical phosphorus concentrations have also been determined by using the same method (Anderson 1993). As diatoms are preserved well in most lake sediments, they can be used to generate long term – series data not only on eutrophication but also from its recovery stages. Predisturbance water chemistry can be determined as targets for lake rehabilitation by examining fossil diatoms (Hall & Smol 1999).
River health can be assessed by using diatoms (e.g. Prygiel & Coste 1999 in France, Harding & Kelly 1999 in UK and Stevenson & Pan in the USA 1999). Diatom assemblages in rivers and streams can be analysed by rigorous statistical techniques to establish their relationship to the environmental factors. The relative abundance of diatom species are used as the most valuable characteristics of diatom assemblages for bioassessment of river health.
An overview of the river health in Western Australia was generated by sampling over 150 sites in the south-west of Western Australia covering an area of 24 000km2 for periphytic diatoms. An artificial substrate collector (The JJ Periphytometer) was used for collecting diatom samples from 46 rivers and streams. Up to 200 species of diatoms were identified in the waterbodies. All sites were classified according to the environmental factors and diatom distribution pattern. This is the largest study of river health assessment conducted in Australia using diatoms at species level (John 1998). The river sites were classified into 4 major groups, the most pristine sites included the streams in native forests in the south-west of Western Australia and the most impacted sites were shown to be affected by eutrophication and salinisation. In between these extremes, there were two groups of sites undergoing varying degrees of degradation. Any new sites sampled for diatoms can be tested to see where they fit in the classification system. Such a system for bioassessment is a valuable management tool.
This publication is part of a similar study involving urban streams and drains in the Perth metropolitan area has been completed recently (John 2000). Identification of diatoms to the species level and enumeration of relative frequency of each species was essential to achieve maximum benefits from the study. Surveys involving generic level of identification of diatoms and recording of mere absence or presence will achieve only limited success (Chessman et al 1999).
In Australia there is large potential for this type of investigation. In a study of sediment analyses of fossil diatoms of an urban wetland in Perth the fossil diatom showed 4 different stages of development in the past (John et al. 1992). The ratio of planktonic diatoms to periphytic diatoms present in the fossil gave an indication of water level changes in the past. Past changes in the nutrient status, salinity and pH as well as climatic changes and sea level changes can be inferred by studying fossil diatoms in the cores extracted from freshwater lakes and oceanic deposits with dates of the deposits determined by radioactive isotope techniques such as 14C and 210Pb, the latter giving fine resolution.
Multivariate calibration methods can be used to develop diatom based transfer functions with which to infer past changes in the salinity of lakes. Lakes are intimately linked to hydrologic and climatic changes (Fritz et al. 1999). Gell (1997; 1998) investigated saline lakes in South Australia by using transfer function.
Climatic changes trigger lake level fluctuations and alters ionic concentrations in water. Diatoms are extremely sensitive to pH and salinity. The distribution of diatom species is clearly related to ionic concentration and composition of the water bodies. The strong correlations between diatoms and ionic concentration enable diatoms to be used to reconstruct past changes in lake water salinity driven by hydrologic and climatic change (Fritz et al. 1993; Fritz et al 1999; Blinn 1995). Environmental changes in marine, brackish waters and estuaries can be inferred by diatom studies, although the techniques and interpretations are more challenging than those used in freshwater lakes and rivers (John 1983a; Snoeijs 1999).
Salinity, nutrients, temperature, rainfall and urbanisation are some of the factors influencing diatom distribution patterns in the estuaries, brackish waters and urban streams. Palaeoecological studies on the environments of fluctuating salinity will be of tremendous importance as diatoms are often the dominant component of the benthic microalgal assemblages in estuarine and shallow coastal environments as anthropogenic input of nutrients pose a severe threat to estuarine and shallow coastal waters (Sullivan 1999).
Diatomite
Fossil deposits of diatoms since the Eocene (about fifty million years ago) in the freshwater environment and since Cretaceous (about eighty million years ago) in the marine environment are mined in different parts of the world. Diatomaceous earth (diatomite) has more than 500 commercial uses (Harwood 1999). The large surface area and porosity provided by diatom frustules and the extremely high melting point of the opaline silica (SiO2 in H20) make diatomaceous earth valuable as a filter, absorbent filler and as an insulating agent. Additionally it is used as an abrasive, pesticide, catalyst in metal polishes, food additive, caking agent, paper and plastic industry and paints.
Lake Gnangara is the largest lake in the Perth metropolitan area. The deposit of diatomite in this wetland was first described by the pioneer geologist Simpson in 1903. Lake Gnangara occupies a shallow circular depression in the quaternary superficial formation with an area of 128ha. In the 1940s the lake was dredged for diatomite which had very high sulphide content. After removing several hundred tonnes of diatomite, the operation was abandoned without any attempt at rehabilitation. Prolonged dry spells and decline in ground water due to land use in the catchment, the water level in the lake started declining in the 1960s. The sediments with disturbed diatomaceous earth containing sulphide were exposed resulting in gradual acidification. In 1976, a pH between 4 and 4.5 were recorded and since then the lake has continued to be acidic with a pH of 3.5 – 4. Currently there are only very few biota in the lake with 6 species of living diatoms where as the diatomite flora was very high with more than 100 species (John unpublished).
Diatoms in Forensic Science
Almost three decades ago, I got interested in diatoms when I was approached by a forensic pathologist to identify the diatoms he retrieved from the lung tissue of a dead body found floating in the Swan River, Western Australia. Application of diatoms in forensic science mostly relate to diagnosis of death by drowning (Peabody 1999). Water enters the lungs and blood stream when drowning. It would be easy to determine whether drowning occurred in a private home bathtub or in the freshwater lake or in an estuary. However diatom based diagnosis may not be all 'smooth sailing'. Lungs, liver, kidney and even brain might harbour diatoms accumulated in a life time especially by a fisherman or sailor who has a long association with rivers, estuaries and marine waters. Diatoms that make their entry into the body over a long period and remain lodged have to be separated from those that get engulfed during drowning. Diatoms can aid in other forensic cases: examination of wet clothing of a suspect may reveal diatoms he or she may have just acquired while rushing through a wetland or garden pond after committing a crime. Good evidence of association may be provided by examination of a variety of materials related to crime scenes, such as paint and metal polish which may contain diatomaceous earth as an ingredient (Peabody 1971; 1977; 1980).
Toxic Diatoms
There is a very small group of species capable of forming harmful blooms and producing toxins in marine waters and estuaries. While the vast majority of diatoms even in blooms are harmless, some are known to cause fish kill, particularly in fish farms, due to oxygen depletion and production of mucilage clogging the fish gills. The tiny but sharp spines on the setae of a few species may injure fish gills and cause their death. In some instances demise of massive bloom may be followed by death of benthic shellfish due to oxygen depletion caused by bacterial decomposition (Fryxell and Villac 1999).
Classification and Systematics
The systematics of diatoms has been almost exclusively based upon frustule characteristics: shape, size, symmetry, structure and density of striae, nature of raphe, copulae and processes on the valves. However, increasingly, there is a trend towards ultrastructure and features of living cells such as chloroplastids, life cycle pattern and reproductive biology to be considered in the systematics. The biological species concept based upon reproductive isolation is difficult to apply to diatoms (Round et al. 1990). It has been shown that a few forms of the pennate freshwater diatom Sellaphora pupila will only breed with the same form whereas others can be induced to interbreed (Mann 1989) under laboratory conditions. Establishing the validity of species on the basis of breeding is laborious and not a pragmatic approach. A natural classification based on the similarity of phenetic characteristics is favoured by Round et al. (1990). Although there is general agreement on the principle of 'a natural classification', in practice there is a vast difference in the interpretation of criteria used by different taxonomists. For example, Lang-Bertalot and his colleagues referred by Mann as "Frankfurt school" have been prolific in their output on taxonomic papers especially description of several new species (Mann 1999). Many of the British diatomists may disagree with the "Frankfurt school's" approach to diatom taxonomy in practice due to inconsistent criteria and 'perceptions of relative weightage to those criteria'. On the other hand, the British scientist Mann and his co-workers themselves have recognised several 'demes' (taxonomically closely related assemblages within a species). Phenotypic plasticity and polymorphism in diatoms become apparent only by examination of large populations of the samples from a locality. An extreme view is represented by a few researchers who believe that the phenotypic plasticity and polymorphism might be attributed to changes brought about during ontogeny , in which case there may be fewer species than currently recognised argues Schmid (1994). Taking species of Sellaphora pupila as a model, Mann and his co-workers through studies of natural population and cultures have held the view that minor variation in shape, size and striation density have a genetic basis truly transmitted by asexual and sexual phases (Mann 1999).
Ideally, variation in valve morphology and anatomy, characters in live forms, physiological and biochemical and molecular and reproductive features should be considered in determining the taxonomy of diatoms. It is almost impossible to combine all these characters together in determining the taxonomic status of a given species. However, just on the basis of valve morphology of a small number of frustules alone, describing 'new species' is currently discouraged. Natural population of large number of a taxon should be studied incorporating as many characters as possible in delineating a species.
The current classification (Round et al. 1990) recognises diatoms as a division:
- Bacillariophyta – with three major classes
- Coscinodiscophyceae – (Centric diatoms)
- Fragilariophyceae – (Araphid – pennate diatoms) and
- Bacillariophyceae – (Raphid – pennate diatoms)
The International Journal of Diatom Research – the official journal of the international society of diatom researchers follow the above classification. The details of the classification is not presented here as I am attempting to give a 'practice guide' based upon light microscopic structure rather than a natural phylogenetic guide.
The centric diatoms, predominantly planktonic include generally radially symmetrical diatoms with no raphe. The araphid pennate diatoms are generally bilaterally symmetrical without any raphe. They are largely epiphytic/epizoic/episamic or periphytic. The raphid pennate diatoms may be symmetrical or asymmetrical, have raphe at least in one valve. In the subclass Eunotiophycidae ("Proraphids") raphes are very short and confined to the poles of the valves. In Achnanthales ("Monoraphids"), a raphe is present in only one valve: the frustule having a raphe valve (RV) and a rapheless valve (RLV) (Fig. 9 a & b). In the rest of the members of Bacillaiophyceae ("Biraphids"), the raphe may be placed longitudinally at the centre (e.g. Naviculales) or laterally shifted to one side (e.g. Cymbellales) (Fig. 5) or eccentrically located on a keel (e.g. Nitzschia) or on the margins of the whole valve (e.g. Surirella) (Figs. 4, d, h, m, n & 7 d).
Diatom Research in Australia
Apart from early occasional records of diatoms in Australia in Schmid's Atlas (1874-1904) and marine diatom studies by Wood (1961a, 1961b, 1963a, 1963b and 1964) and Wood et al. (1959), diatom study was sparse in Australia, particularly on inland diatoms. An illustrated species list with information on water quality preferences from Eastern Australia by Foged (1978) and similar studies on diatoms of the Alligator Rivers Region with emphasis on ecology by Thomas (1983) heralded the modern era of diatom research in Australia. The first systematic treatise illustrated with micrographs, taxa descriptions and ecological information on diatoms in Australia was by John (1983a) who covered 360 taxa of the Swan River Estuary, Western Australia and included both marine and freshwater species. John, over the past 25 years of diatom research, has described 25 species of diatoms (inland and marine) new to science from Western Australia. Within the last 20 years several new genera and species were also described from other parts of Australia, particularly Tasmania (e.g. Blaze 1984; Hayworth & Tyler 1993; Vyverman et al. 1997, 1998).
Ultrastructure and distribution patterns of several inland and marine species of diatoms have been investigated by Australian diatomologists e.g. (John 1986, 1993a; Hallegraeff 1992). Recently John has completed research projects on the use of diatoms as tools for assessing health of rivers and streams in the south west of Western Australia with iconographs of diatoms (John 1998). We have also examples of palaeoecological studies using diatoms to infer past environmental conditions (John 1993c; Gell 1997, 1998; McBride & Selkirk 1998). It is extremely difficult to arrive at the exact number of diatom species recorded from Australia, because of the impossible task of validating many of the records with no micrographs, diagrams or voucher slides or good descriptions. The International Diatom Herbarium at Curtin University, Perth, WA, has a collection of over 20, 000 slides currently being catalogued and curated. In a bibliographic checklist of non-marine algae in Australia (Day et al. 1995) 828 species and 381 infraspecies of diatoms are mentioned. It will be very challenging to follow the trails of diatom records in this publication and verify their validity. The significance of diatom research in Australia was recognised when Australia hosted the 15th International Diatom Conference in 1998 in Perth, Western Australia.
Although the diatom flora and biogeography of diatoms in Australia have not been adequately explored, a sound foundation has been laid for future work on urban streams by the current project.
Chapter 3 Features & Illustrated Glossary for Diatom Identification
- Glossary for Diatom
- glossary applicable to urban stream diatoms
Identification of diatoms to the generic and species level requires an understanding of the basic morphology of cells explained in the previous chapter. The following is a short list of criteria generally used for the classification of diatoms. The features required for identification can be observed under Light Microscopy. Most of the features can be seen under 40X magnification, but Oil immersion (100X) is recommended for observation of striae and areolae, and raphe endings (fissures).
- Habit of cells single or colonies (Fig. 2).
- Shape and size of frustules, and valves, symmetry, girdle bands (Copulae) (Figs. 1, 4, 5, 10 & 11).
- The number, shape and position of chloroplastid in the cell (Fig. 7). Diatoms have to be freshly collected or preserved in Lugol's solution for observation of plastids, which provides additional information for identification. Examination of fresh or preserved sample may not be always possible and not essential for identification. However, plastids do provide helpful additional information for confirmation of identity. As one becomes experienced, live examination does provide a method of 'rapid identification', to the generic level and even species level for a few taxa.
- The ends of valve shapes (poles): cuneate, rounded, rostrate, capitate (Figs. 10 & 11).
- Raphe slits, structure: simple, broad filament, twisted or straight, central and terminal endings (fissures), position, whether on a keel wing, embedded in siliceous ridge or located on the perimeter of valve (Figs. 7 d & 12).
- Axial area: sternum shape, dimensions, whether thickened or not (Figs. 12 & 13).
- Central area and central nodule, shape, size, thickened, expanded or inflated (Figs. 12 & 13).
- Polar nodules: shape, size, hyaline or with markings (Figs. 12 & 13).
- Girdle bands (Copulae) extensions of bands: Septa and Partecta (Chambers) (Figs. 1 & 4).
- Striae: structure: pores, areolae, alveoli, position, orientation (parallel, radiate). Striae density i.e. number of striae per 10 µm (Figs. 12 & 13).
- Hyaline areas (clear silicified areas) on valve face (e.g. Lyrate).
The glossary of the basic terms used in the description of genera and illustrations on the above features are given below.
Glossary Applicable to Urban Stream Diatoms
- Acicular = needle shaped
- Alveoli = Transversely linear chambers in the valve with a small opening externally and large opening internally
- Apex = pole (end) of diatom valve
- Apical axis = axis connecting the two poles of a valve
- Apiculate = pole (apex) of valve tapering to a point
- Araphid = diatoms without any raphe
- Arcuate = strongly curved
- Areolae = chambered pores in the diatom valve
- Axial area = the area between the raphe and striae on the valve face
- Bifurcate = divided into two branches
- Biraphids = Diatoms with raphe on both valves
- Biseriate = two rows of pores or areolae
- Canal raphe = raphe lying in a distinct groove
- Capitate = pole (end) swollen
- Central area = the clear area at the centre of the valve face
- Central ending = the external groove of the raphe slit at the centre of the valve
- Central nodule = the thickly silicified area in between the central endings of the raphe slits
- Cingulum = girdle
- Clavate = club-shaped
- Convergent striae = striae converging (angling) towards the poles
- Copulae = girdle bands
- Costae = distinctly thickened ribs on the valve face
- Cruciform = cross-shaped
- Cuneate = wedge-shaped
- Cymbelloid = dorsiventral valve with raphe close to the ventral margin
- Dorsal margin = the convex margin
- Dorsiventral = valve with one convex, and one straight or concave margin
- Eccentric raphe = raphe deviating from the apical axis
- Elliptical = shape of valve ellipsoid
- Epitheca = the larger overlapping valve
- Fascia = a distinct central area extending to the margins of the valve, without any marking
- Fibula(e) = Siliceous bridge on either side of a raphe
- Foot pole = narrow half (basal) of a club-shaped valve
- Frustule = the cell walls of a diatom connected by girdle bands
- Girdle view = Side view of cell with girdle bands
- Head pole = broad (upper) half of a club-shaped valve
- Heteropolar = poles (apices) of valve being dissimilar
- Hyaline area = Clear area (siliceous) without any marking
- Hypotheca = smaller underlying valve
- Isopolar = poles of a valve being similar shape and size
- Keel wing = a raised part of a valve face eccentrically located to the apical axis
- Lanceolate = lance shaped
- Lenticular = lens shaped
- Linear = long and narrow almost uniform in width
- Linear-clavate = elongated club-shaped
- Linear-elliptic = elongated with inflated ellipsoid margins
- Longitudinal striae = striae parallel to the longitudinal axis (apical axis) of the valve
- Lunate = crescent shaped
- Mantle = the side (outermost) part of valve bending downwards from the surface to the girdle
- Monoraphid = Diatom with a raphe only on one valve
- Mucilage = viscous polysaccharide secreted by diatoms
- Multiseriate = many rows of pores or areolae
- Orbicular = spherical
- Ovate = oval shaped
- Panduriform = shaped like a fiddle ('8' shaped)
- Parallel striae = striae arranged more or less parallel to each other
- Pennate = valve with laterally symmetrical
- Polar nodule = thickened silicified area of the pole where the terminal ends of the raphe are located
- Proraphid = Diatoms with a small raphe at the poles
- Puncta = poroid opening of the siliceous valve – a traditional term for areolae
- Radiate striae = striae pointing to the centre of the valve
- Raphe = a longitudinal slit (groove) in the valve face located between the two polar nodules, with two raphe branches
- Raphe branch = refers to the raphe branch from the centre to the pole. A raphe consists of two raphe branches
- Raphe-sternum = thickened hyaline axial area in pennate diatom (pseudoraphe)
- Rostrate = the apex of a valve ending like a beak
- Septum = the inward expanded plate from the valvocopula (girdle band)
- Sigmoid = shaped like 'S' with the ends curving in opposite direction
- Stauros = thickened central nodule extending to the margin of valve
- Stellate = star shaped
- Stigma(ta) = an isolated pore different from the pores of the striae usually located lateral to the central area
- Stria(e) = row of pores in valve
- Terminal endings (terminal fissures) = polar endings of raphe
- Transapical axis = axis through the centre connecting the margins across
- Transverse striae = striae running across the valve faceUndulate = wavy
- Uniseriate = single row of pores or areolae
- Valve face = the surface of valve surrounded by the mantle
- Valve margin = the edge of valve face
- Valvocopula = copula (girdle element) closest to the valve – different in structure from the rest of the cingulum
- Ventral = margin (straight or concave) opposite to the dorsal side
| Fig 10 Shapes of valves and poles and raphe position |
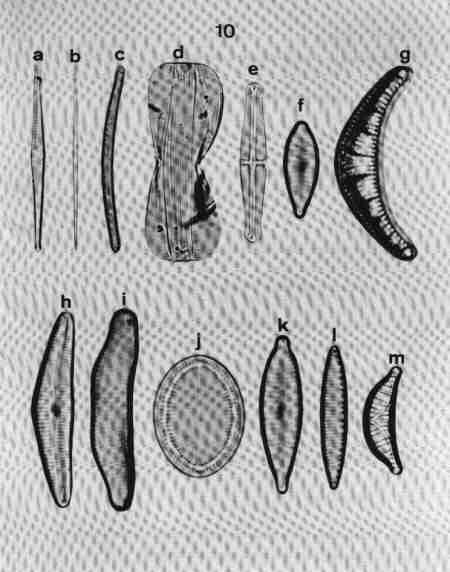
(original size image) |
- Linear rostrate pole with 'fascia' (hyaline central area reaching margins).
- Acicular (needle shaped).
- Arcuate with capitate poles.
- Bilobate with raphe on keel wings.
- Lanceolate with capitate ends.
- Clavate with rostrate ends, Heteropolar (Head pole different from foot pole).
- Arcuate and valve surface costate (costae across).
- Dorsiventral with rounded ends.
- Dorsiventral and arcuate.
- Elliptic.
- Heteropolar (dissimilar poles) with rostrate poles.
- Keel on the one margin – with fibulae and keel puncta.
- Arcuate, dorsiventral – costate, arched keel with raphe.
|
| Fig 11 Morphology – Shape of valves and poles |
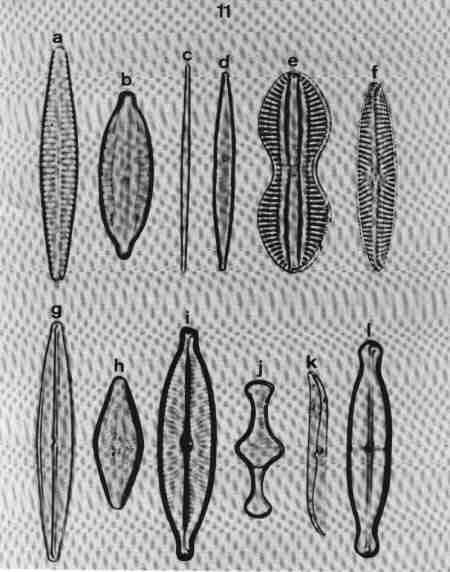
(original size image) |
- Lanceolate and parallel striae; narrow axial area.
- Broadly lanceolate area with rostrate poles.
- Linear – narrow – acicular.
- Linear – lanceolate.
- Panduriform; raphe flanked by siliceous ribs.
- Lanceolate with radiate striae towards centre.
- Lanceolate with rounded ends, raphe in siliceous ridge; raphe endings surrounded by siliceous ridge, like a pencil – point.
- Rhombic shape.
- Lanceolate with slightly produced rostrate ends.
- Face view of valvocopula with a septum.
- Sigmoid valve.
- Lanceolate valve with capitate ends; a central 'stauros' (siliceous thickening from the central area to the margins).
|
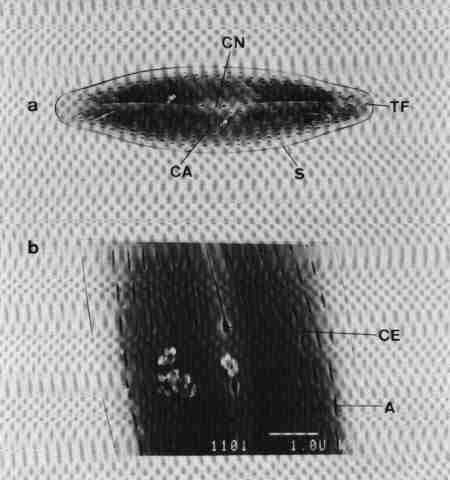
| Fig 12 A typical pennate diatom 'Pinnularia' |

(original size image) |
|
TN = Terminal nodule (polar nodule)
TF = Terminal fissure (external ending of raphe)
AA = Axial area
CE = Central ending of raphe slit
CA = Central area
CN = Central nodule (thickened with silica)
R = Raphe: curved and filamentous in this genus
S = Striae – made of alveoli
Striae density = Number of striae per 10(m normally measured from the central area upwards or downwards
In this diatom Pinnularia the striae are made of ALVEOLI (elongate chambers extending from the axial area to the periphery. There may be several tiny pores (areolae) on the outer surface and an elongated channel on the lower surface.
|
| Fig. 13 SEM structure of a typical Pennate Naviculoid diatom |
(original size image) |
- Valve view
CA = Central area
CN = Central nodule
S = Uniseriate striae – consists of apically linear pores (areolae)
TF = Terminal fissure (terminal ending), note the curving
- Enlarged view of the central part of valve
C E = Central ending (an expanded pore)
A= Areolae – opening outside through linear pores
|
Chapter 4 Short Diagnostic Descriptions of Diatom Genera in Urban Streams
- short diagnostic descriptions of diatom genera in urban streams
- I Centric Valves circular (radially symmetrical), no raphe many discoid or lobed plastids
- II Pennate diatoms Valves elongate, with or without raphe
I CENTRIC
Valves circular (radially symmetrical), no raphe, many discoid or lobed plastids.
1. Aulacoseira (Fig. 14 a)
Frustules in girdle view, cylindrical, linked to form long chains, mostly lying in girdle view. Valve mantle with areolae arranged in straight or decussate (curved) rows. Spines present around the circular margin, valve face uniformly areolate. Species of the genus mostly in mesotrophic or eutrophic but rarely in oligotrophic waterbodies.
2. Cyclotella (Fig. 14 b)
Frustule in girdle view, drum-shaped, valve surface circular, undulate or flat, demarcated into a marginal zone of alveoli with no conspicuous marginal spines, and a central area with irregular markings. Cells mostly seen in valve view. Found mostly in mesotrophic waterbodies. Typically planktonic, but often found in benthic form in shallow water bodies of high conductivity.
3. Melosira (Fig. 14 c)
Frustules in girdle view, cylindrical, united in distinct pairs to form filaments. Valve face flat with small areolae. Species of Melosira are found in oligotrophic to eutrophic, fresh, brackish and marine waters.
4. Paralia (Fig. 14 d)
Cells linked to form chains; plastids discoid, many. Frustule disk-shaped. Valve mantle coarsely furrowed and ornamented, furnished with robust locules. Valve face circular with hyaline central area and coarse costate radial markings or dots.
This was described as Melosira sulcata but transferred to Paralia and is the most common benthic diatom in estuaries of Western Australia. It was observed in streams close to the estuary influenced by salinity.
5. Skeletonema (Fig. 14 e)
Frustules discoid or spherical, in chains connected by marginal processes, seen invariably in girdle view, like short beads joined by several threads. It is an estuarine and marine planktonic species, but seen in brackish waters under marine influence.
6. Thalassiosira (Fig. 14 f)
Frustules in girdle view, drum-shaped, delicate, thin, solitary or in chain joined by central mucilaginous threads. Valve face circular, flat or undulate with visible marginal spines and processes, fine areolae, mostly radial (see Fig. 6 for details of the morphology). Observed in marine brackish waters; often seen in valve view.
| Fig. 14 |
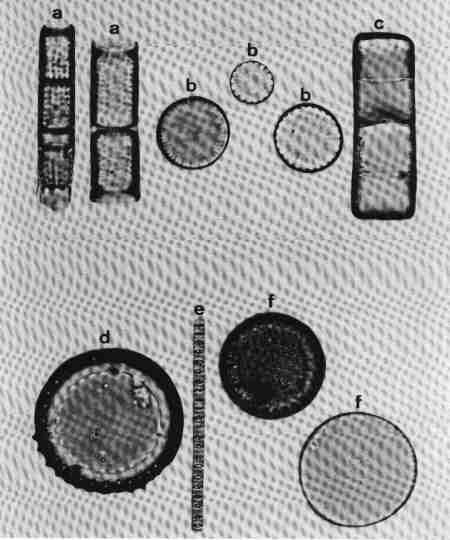
(original size image) |
|
Diatom Genera
Centric diatoms
- Aulacoseira (girdle view)
- Cyclotella (valve view)
- Melosira (girdle view)
- Paralia (valve view)
- Skeletonema (girdle view of chain of cells)
- Thalassiosira (valve view)
|
II PENNATE DIATOMS
Valves elongate, with or without raphe.
A. ARAPHID GROUP
Pennate diatoms without raphe but with a clear axial area.
1. Ctenophora (Fig. 15 a)
Cells narrow, needle-like. Valve flat, linear with rounded or slightly capitate ends; epiphyte, attached by a mucilage pad to substrate; two plate like plastids present. Striae transverse, parallel, in single rows of rectangular areolae. Axial area narrow with a thick hyaline central area (fascia). Areolae tend to be smaller towards the axial area. There is an apical pore field. This is a freshwater genus tolerant of eutrophic conditions and can be distinguished from Synedra by its fascia and areolae.
2. Fragilaria (Fig. 15 b)
Frustules joined to form ribbon-like colonies linked through marginal processes; two plastids. Valve linear, linear-lanceolate, elliptical, cruciform or triangular, poles capitate, rounded or rostrate. Valve face areolate; areolae arranged in transverse rows. This is predominantly a freshwater genus and resembles Staurosira very closely, but has striae composed of simple pores and areola are also present in copulae (girdle bands).
3. Martyana (Fig. 15 c)
Valves, ovate, elliptical, rarely heteropolar, striae coarse, made of single row of large slit-like areolae with a narrow axial area. This genus was separated by Round (1990) from Opephora based on the description of Opephora martyi (Opephora is a marine genus). Opephora is clavate, markedly heteropolar with larger axial area. Martyana can be distinguished from Opephora by its narrow axial area. This genus is rare in Perth urban streams.
4. Meridion (Fig. 15 d)
Valves distinctly heteropolar and clavate with rounded 'head pole' and rostrate or capitate "footpole". Frustules forming fan shaped colonies attached by their valve face. The valve is crossed by transapical costae, striae fine, parallel consisting of simple pores. Axial area narrow. It is a freshwater genus, rare in urban streams.
5. Staurosira (Fig. 15 e)
Frustules single or in straight colonies or zig-zag filaments; a pair of plastids in each cell. Valves oval, elliptical or cruciform, with widely spaced uniseriate rows of circular or elliptical areolae. Common in shallow freshwater streams.
6. Synedra (Fig. 15 f)
Frustules, elongate, narrow. Valve linear, isopolar, lanceolate with rounded or capitate ends. Axial area narrow – some species have a clear transverse central area. Striae parallel, with simple areolate structure in single rows - continuing on to the mantle. Pore field present at each pole. This is an exclusive freshwater genus, epiphytic or periphytic in habit and differs from Tabularia in striae and axial area.
7. Tabellaria (Fig. 15 g)
Cells arranged in the form of zig-zag chains connected to each other at the corners, each with many strip-like plastids.
Frustules seen mostly in girdle view, characterised by several girdle bands. Valve linear, isopolar, inflated in the middle and poles; striae fine, parallel – made of single row of areolae. There is an isolated pore located lateral to the centre. This is a very distinct genus easy to identify and found in slightly acidic waterbodies with low concentration of ions and indicative of pristine streams.
8. Tabularia (Fig. 15 h)
Frustules needle-like, single, or in clusters, epiphytic or periphytic. Valve linear, isopolar with capitate ends; striae transverse short with two rows of areolae arranged towards the margin leaving a wide axial area. There is a distinct 'porefield' at each end. Tabularia differs from Synedra in the structure of striae and axial area. This is predominantly a marine or estuarine genus but found in brackish waterbodies.
9. Thalassionema (Fig. 15 i)
Valves elongate, linear, isopolar, forming stellate, zig-zag colonies; plastids discoid. Ends of valves slightly tapering and rounded. Valve face flat with large axial area, areolae reduced and confined to the margins. This is a marine/estuarine genus and may be found in brackish part of the estuary, but rare.
| Fig. 15 |
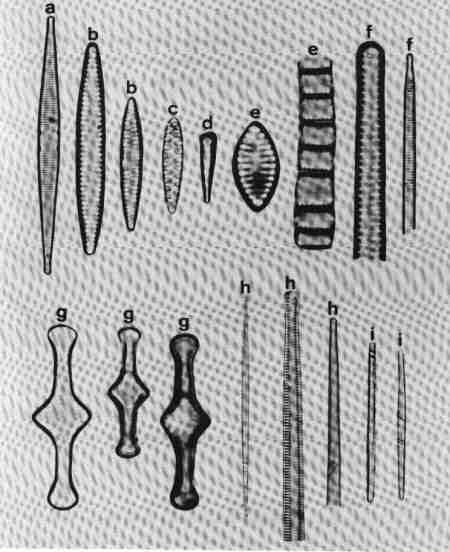
(original size image) |
|
Diatom Genera
Pennate – Araphids (genera without raphe)
- Ctenophora - valve view
- Fragilaria - valve view
- Martyana - valve view
- Meridion - valve view
- Staurosira - valve and girdle views
- Synedra - valve view
- Tabellaria - valve view
- Tabularia - valve view
- Thalassionema - valve view
|
B. MONORAPHID GROUP
Diatoms in this group are Heterovalvar (two valves with different structure): raphe present only in one valve; the two valves are referred to as Raphe Valve (RV) and Raphe Less Valve (RLV). This group is very common in urban streams.
1. Achnanthes (Fig. 16 a)
Cells may be single or in colonies attached to the substrate by mucilage stalks (see Fig. 2 b). Valves, linear or linear-lanceolate, frustules slightly bent in the middle; plastids many. Areolae complex, separated by robust costae. This is predominantly a marine or brackish water species but rarely found in urban streams due to the estuarine influence.
2. Achnanthidium (Fig. 16 b)
Frustules attached to macrophytes or substrates. Valves linear, linear-lanceolate, or elliptical with rounded or capitate ends and delicate; RV and RLV have usually the same striae structure: uniseriate (consisting of single rows of pores) or multiseriate (many rows of pores). Achnanthidium species are generally less robust than 'Achnanthes' species. Axial area narrow, linear, or lanceolate with expanded central area. The members of this group were placed previously under Achnanthes. Species of Achnanthidium are very common in urban streams.
3. Cocconeis (Fig. 16 c)
Cells, epiphytic or periphytic with one flat plastid. Valves elliptic or circular. RV and RLV with distinctly different striae. Axial area narrow. Single rows of elongate or circular pores forming transverse regular or irregular rows of striae. Raphe simple, no terminal tissues. A common genus in almost all streams.
C. PRORAPHID GROUP
Diatoms with short raphe present only at the poles and seen clearly only in girdle view and not in valve view are included in this group. The cells are basically dorsivental, isopolar, and not bilaterally symmetrical.
1. Eunotia (Fig. 16 d)
Cells single or in colonies; in girdle view rectangular. Valves dorsiventral, arcuate, slender, and linear with rounded or capitate ends; dorsal margin convex, ventral margin, straight or concave. A small raphe at the poles visible only in girdle view. Common in pristine sites.
| Fig. 16 |
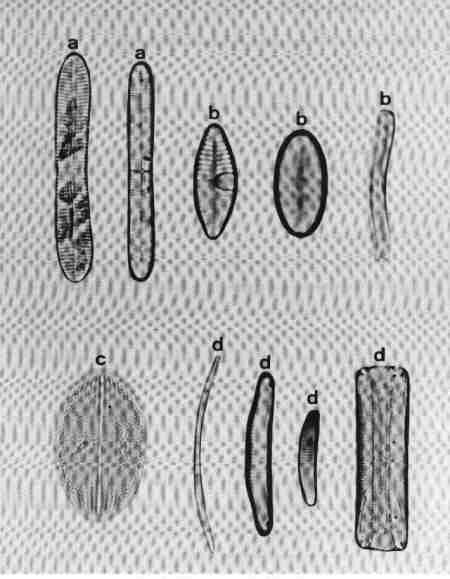
(original size image) |
|
Diatom genera
Monoraphids (genera with raphe on one valve)
a) Achnanthes RLV valve
RV valve
b) Achnanthidium
- valve view
- valve view
- girdle view
c) Cocconeis RLV - valve view
Diatom genera - Proraphid
d) Eunotia
- valve view
- valve view
- valve view
- girdle view
(short raphe can be seen clearly)
|
D. BIRAPHID GROUP
These include all diatoms with both valves containing raphe.
D.1 NAVICULOID
Bilaterally symmetrical, isopolar valve boat shaped with two raphe slits, extending from each pole to the centre with central and terminal fissures. Raphe occupying the apical axis of the valve surrounded by the axial area. This is the largest group of fresh water diatoms. Raphe not on a keel.
1. Aneumastus (Fig. 17 a)
Cells single, two plastids one on either side of the median transapical plane. Valve lanceolate with rostrate or capitate ends, valve face flat. Axial area narrow. Central raphe endings expanded, terminal fissures curved to opposite poles. Central area elliptical, expanded. Striae made of single row of pores, parallel; pores smaller towards the edges of valve. Previously placed under Navicula and then Mastogloia, Aneumastus was erected as a separate genus by Mann and Stickle (Round et al. 1990). Rarely found in urban streams.
2. Anomoeoneis (Fig. 17 b)
Cells with single highly lobed plastid. Valve elliptical, lanceolate with capitate or rostrate ends; striae arranged in longitudinal rows of single oval areolae in irregular undulating (wavy) manner; valve face free of areolae towards the middle of the valve. Axial area narrow bordered by a single row of areolae. Central area irregular expanding almost to the margin unilaterally. Raphe slits extends to the very end of the poles with hooked terminal fissures. This genus is easy to identify due to the 'lyrate' clear area on the valve face. It is found in high conductivity water.
3. Brachysira (Fig. 17 c)
Cells, single, one plastid. Valve linear-lanceolate or rhombic, striae made of transapically elongated pores in undulating vertical rows. Axial area narrow but well silicified. Raphe ends are simple and short.
This genus was previously placed in Anomoeoneis (Round et al.1990). This is an epipelic genus characteristic of oligotrophic streams and may be present in pristine sites. Some species are found in saline waterbodies.
4. Caloneis (Fig. 17 d)
Valves, cylindrical, linear or elliptic-lanceolate with rounded or capitate ends, inflated in the middle in a few species. Raphe straight, axial area narrow, central area round or elliptic like a fascia, rarely with a pair of lunate markings. Raphe straight, simple. Striae parallel made of fine alveoli longitudinally intercepted by hyaline bands closer to the margin. (These bands are inner openings of the alveoli). The striae have a costae-like look. This genus is distributed in both fresh and brackish waters.
5. Craticula (Fig. 17 e)
Cells with two elongate plate like plastids. Valves lanceolate with capitate or rostrate poles, striae parallel made of single rows of pores arranged longitudinally and transversely. Areolae complex in structure. The diatom is characterised by irregular large transverse costae across the valve. Formerly included under Navicula and has been elevated to new genus level (Round et al. 1990). This is an epipelic genus preferring the brackish water or water of high conductivity.
6. Diadesmis (Fig. 17 f)
Cells in chain connected by interdigital structures at the margin of the valve; single plastid. Valves linear, lanceolate or elliptic-lanceolate; valve face flat. Striae discontinuous towards the central area, made of single row of transapically elongate pores. Raphe, simple not curved. Central raphe endings distantly arranged. Axial area widening to a broad lanceolate area. This used to be under Navicula and the description of the genus is based upon Navicula confervacea (Round et al.. 1990). It is a freshwater genus of subaerial habitat. Commonly found attached to concrete drains in urban streams.
7. Diploneis (Fig. 17 g)
Valves elliptic to linear or constricted in the middle (panduriform) with broadly rounded endings. The raphe is flanked by distinct siliceous ridges; axial area and central area raised and bounded by longitudinal canals. Striae alveolate, uniseriate or biseriate; separated by transverse costae. Polar modules prominent. This is a distinct genus found mostly in brackish or high conductivity waterbodies.
8. Fallacia (Fig. 17 h)
Cells with a single H shaped plastid with two girdle oppressed plates. Valves delicate, linear, lanceolate or elliptical with bluntly rounded ends. Striae consists of a single row of areolae, parallel interrupted by a lateral lyrate region (the central area joining the hyaline lateral area on each side of the valve face forming the lyre shaped configuration). Central endings of the raphe expanded into pores. External terminal fissures curved at the very end of poles. This is a large genus with both marine and freshwater species and epipelic. This genus was separated from Navicula by Mann and Stickle (Round et al.. 1990) based on the description of Navicula pygmaea.
9. Frustulia (Fig. 17 i)
Cells single, occasionally grows in mucilage tubes, seen mostly in valve view. One H shaped plastid. Valves linear-lanceolate, rhombic or linear elliptic. Striae made of fine pores. Raphe straight, flanked by prominent siliceous ribs, the terminal raphe endings on ridges looks like a 'pencil point' and located same distance below the pole. This is an easy genus for identification due to the 'pencil point' raphe endings and prominent siliceous raphe sternum.
| Fig. 17 |

(original size image) |
|
Diatom Genera
Biraphids – Naviculoid
- Aneumastus – valve view
- Anomoeoneis – valve view
- Brachysira – valve view
- Caloneis – valve view
- Craticula – valve view
- Diadesmis – valve view
- Diploneis – valve view
- Fallacia – valve view
- Frustulia – valve view
|
10. Hyppodonta (Fig. 18 a)
Cells single with two plastids arranged in a staggered manner. Valve linear, lanceolate with rounded, rostrate or capitate ends; axial area very narrow, central area irregular. Poles devoid of striae, characterised by a thick clear hyaline area. Striae made of transapically linear coarse areolae. This is a freshwater genus and tolerates a wide range of conductivity. This was separated from Navicula by Lang-Bertalot (1996). The description of the genus is based on Navicula capitata.
11. Luticola (Fig. 18 b)
Cells single, one plastid, usually seen in valve view. Valves linear, lanceolate or elliptical with blunt rounded or capitate ends. Axial area narrow, central area often widened into a transverse stauros with a distinct isolated pore (stigma). Striae slightly radiate with clear simple distantly arranged pores, irregularly shortened at the centre. Terminal and central raphe ending slightly curved.
This is common in fresh or brackish water streams with erosion problem; it is also found in subaerial or aerial habitat. This was separated from Navicula by Mann (Round et al.. 1990), due to the structure of stigma, stauros, and raphe. The genus description is predominantly based on Navicula mutica.
12. Mastogloia (Fig. 18 c)
Cells single with two plastids – each placed towards the pole. Valves lanceolate, elliptic, rostrate or capitate. Striae of single or two rows of areolae in some species, interrupted by a lyrate hyaline area. Central raphe endings expanded, and terminal fissures hooked. The girdle band (valvocopula) close to the valve with chambers (Partecta – marginal locules).
This genus is easy to identify because of the marginal locules (Partecta) like a pair of 'dentures' seen clearly in both valves and girdle view. This genus is rare in freshwater bodies but common in brackish, estuarine marine and saltwater bodies and a good indicator of salinization in inland waterbodies.
13. Navicula (Fig. 18 d)
Cells single with two plastids. Valve linear, or linear-lanceolate with rostrate or capitate or blunt ends. Striae arranged transversely in single or double rows of areolae, parallel or radiate. Raphe endings hooked at the poles; axial area narrow.
This is a highly variable genus, several species are being separated from Navicula on the basis of detailed morphological studies into newly created or resurrected genera. Predominantly a benthic genus (epipelic, epilithic or epiphytic) with maximum species in freshwater bodies.
14. Neidium (Fig. 18 e)
Valves single, linear to lanceolate with blunt, rostrate, produced ends; striae in a single row of round and elongated pores. One or more longitudinal hyaline lines are present close to the margin. In some species central raphe endings are bent in opposite directions, almost at right angles, terminal fissures forked, central area large and transverse.
A distinct genus easily identified due to the marginal hyaline lines, broad central area and bent central endings of the raphe. This is an exclusive freshwater genus characteristic of pristine streams, but never found in dominant frequency.
15. Pinnularia (Fig. 18 f)
Cells with two girdle appressed plate-like plastids with highly dissected margins. Valves linear, cylindrical, lanceolate or elliptic with rounded to capitate or rostrate poles. Striae prominent, made of alveoli (chambered with poroid opening externally and elongate apertures internally). Axial area prominent. Raphe with expanded central endings and long hooked terminal fissures. Central area varies; expanding to the edges in a few species. This is a distinct freshwater genus and a good indicator of slightly acidic or humic and relatively pristine, low conductivity urban streams.
16. Sellaphora (Fig. 18 g)
Cells single with H shaped plastid, valve linear, lanceolate or elliptical with blunt or rounded or rarely capitate ends. Striae in single rows of simple areolae. Striae may be parallel, radiate towards the centre. Axial area prominent, extending to the poles with a T shaped siliceous plate. Raphe straight. Central area prominent extending to the margin (stauros) in some species.
This was separated from Navicula due to the structure of plastid, areolae and prominent axial area (Raphe sternum). A typical freshwater genus and epipelic.
17. Stauroneis (Fig. 18 h)
Cell single, two plastids. Valve lanceolate to lanceolate-elliptic with rounded or rostrate to capitate poles. Axial area very narrow. Central area characterised by a narrow transverse thickened fascia (stauros). Striae parallel made of closely arranged poroids. An epipelic freshwater genus.
| Fig. 18 |

(original size image) |
|
Naviculoid Genera
- Hyppodonta – valve view
- Luticola - valve view
- Mastogloia - valve view showing striae, - valve view with marginal partecta of valvocopula
- Navicula – valve view, - valve view
- Neidium – valve view
- Pinnularia – valve view, - valve view
- Sellaphora – valve view, - valve view
- Stauroneis – valve view
|
D.2 SIGMOID GROUP
Sigmoid valves, axial area and raphe, located at the apical axis.
1. Gyrosigma (Fig. 19 a)
Cells with two large plastids – one on each side of the girdle margins – lobed or dissected. Valve sigmoid – linear to lanceolate with rounded poles, striae arranged in longitudinal and transverse rows of areolae at right angle. Axial area narrow, central area small, irregular. Raphe sigmoid, terminal fissures curved to the same direction as the poles; central fissures slightly curved in opposite directions.
This is a distinct genus, the sigmoid valves and the arrangement of the striae are the most important diagnostic features. It is an epipelic genus common in freshwater with high conductivity, common in urban streams.
2. Pleurosigma (Fig. 19 b)
Cells single, two or four ribbon shaped convoluted plastids extending up to the poles. Valves linear to lanceolate, or rhombic-lanceolate with rounded or tapering acute poles. Axial area narrow, central area small. Raphe sigmoid, terminal fissures hooked to opposite sides; central raphe endings expanded and turned to the same side. Striae arranged in transverse and diagonal (oblique) rows.
This genus is easy to identify due to its distinct shape combined with the transverse and oblique arrangement of areolae. Epipelic in habitat, it is a marine and brackish water genus occurring in freshwater of high conductivity, common in urban streams.
D.3 CLAVATE – HETEROPOLAR GROUP
Valve heteropolar and Raphe at apical axis.
1. Gomphonema (Fig. 19 c)
Cells colonial with branched mucilaginous stalk – one plastid lying on one side of the girdle extending beneath both valves.
Valves clavate (club shaped), heteropolar (head pole broader than a foot pole) with capitate to apiculate ends. Raphe central with central endings expanded and terminal fissures slightly curved. Axial area narrow; striae radial to parallel. Stigma often present lateral to the central area. A freshwater epiphytic genus common in most of the streams.
D.4 DORSIVENTRAL VALVES WITH ECCENTRIC RAPHE WITHOUT KEEL WING (Cymbelloid)
1. Amphora (Fig. 19 d)
Cells single, elliptic, in girdle view with one or two plastids. Valves markedly dorsiventral bilaterally asymmetric (Cymbelloid symmetry), semilanceolate, semicircular or arcuate, isopolar with rounded, rostrate or capitate poles. Raphe area narrow. Striae longer on the dorsal side and shorter or absent on the ventral side of the valve, striae made of uniseriate or biseriate areolae. Axial system on both valves seen when the frustule is viewed from the ventral side; girdle bands and parts of the dorsal side of the valves visible when viewed from the dorsal aspect of the frustule. An epiphytic or periphytic genus very common in brackish water bodies and streams with high conductivity, although it is predominantly a marine genus.
2. Cymbella (Fig. 19 e)
Cells solitary or colonial with mucilaginous stalks, one plastid with two H shaped valve appressed plates. Valves lanceolate or arcuate, isopolar, bilaterally asymmetrical (cymbelloied) with rounded, rostrate or capitate poles, raphe eccentric arranged towards the ventral side, terminal endings curved to the dorsal side, central endings expanded, axial area narrow, striae slightly radiate; isolated stigma may be present lateral to the central area. Striae-uniseriate, made of poroids. There is an apical pore field at each pole (clusters of tiny pores at the pole). This is a freshwater genus, epiphytic, epilithic or epipelic common in urban streams especially streams with macrophytes.
3. Encyonema (Fig. 19 f)
Cells with one plastid lying with its centre against the ventral side. Valve with strong convex dorsal margin and straight or concave ventral margins (cymbelloid symmetry); poles acutely or bluntly rounded; dorsal mantle deeper than the ventral mantle; raphe parallel to ventral margin. Striae uniseriate made of longitudinally elongated slits, terminal fissures strongly curved towards ventral margin, well before reaching the poles, no apical pore field.
This genus is very similar to Cymbella and was separated from Cymbella on the basis of orientation of raphe, structure of striae and plastids (Round et al.. 1990). Encyonema has no pore field; the terminal raphe endings are curved ventrally.
D.5 DIATOMS WITH RAPHE RAISED ON KEEL
Valve twisted, raphe on sigmoid keel wing
1. Entomoneis (Fig. 20 a)
Cells single twisted about the apical axis, mostly seen in girdle view in bilobate shape (see Fig. 10 d). There is one or two axial plate-like plastids. Valve linear or lanceolate centrally constructed with acute poles compressed laterally, thinly siliceous and delicate. A high narrow keel (wing) located on the narrow axial area carrying the sigmoid raphe. Striae fine made of tiny pores.
The raphe is difficult to see because of the twisted nature of the apical axis. There may be transverse ribs on the 'wings' of the valve. Several thin girdle bands present. This is a complex genus but with unique and unambiguous characters. Epipelic in habit, it is a marine, brackish genus with very few freshwater species.
Valves with transverse costae
1. Denticula (Fig. 20 b )
Frustule in girdle view, linear or rectangular. Valve view, linear, linear-lanceolate or linear elliptic with rounded poles. Raphe straight, eccentric, transapical costae prominent, striae, fine, made of small poroids. This is a freshwater genus and prefers high conductivity. Rare in urban streams.
2. Epithemia (Fig. 20 c)
Cells solitary, with one large plate-like plastid with highly lobed margin. Valves dorsiventral; or arcuate with blunt or broadly capitate poles. Striae arranged in between robust costae. Raphe eccentric, biarcuate with simple central and terminal endings. Raphe crossed by silica struts on a raised keel; giving a beaded appearance. Central area narrow, no axial area. This genus is epiphytic or periphytic and is common in alkaline streams of high conductivity.
3. Rhopalodia (Fig. 20 d)
Cells solitary, one plate-like plastid. Valve dorsiventral, lunate or kidney shaped, with cuneate or capitate ends, isopolar. Raphe located along the dorsal margin, raised on a keel. This is a distinct genus, epiphytic or periphytic common in high conductivity water. Axial area narrow, prominent transapical costae, with two or more rows of complex alveoli in between. External terminal raphe endings expanded.
| Fig. 19 |
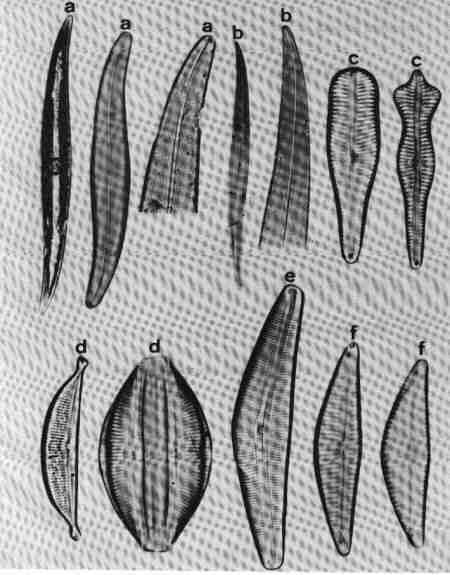
(original size image) |
|
Naviculoid Genera
a) Gyrosigma – fresh cell with chloroplast (valve view)
- valve view
- part of valve showing the horizontal and vertical striae
b) Pleurosigma – valve view, - part of valve showing the oblique striae
Heteropolar – Clavate
c) Gomphonema – valve view, - valve view
Dorsiventral Valves
d) Amphora – single valve view, - frustule
e) Cymbella – valve view
f) Encyonema – valve view
|
| Fig. 20 |
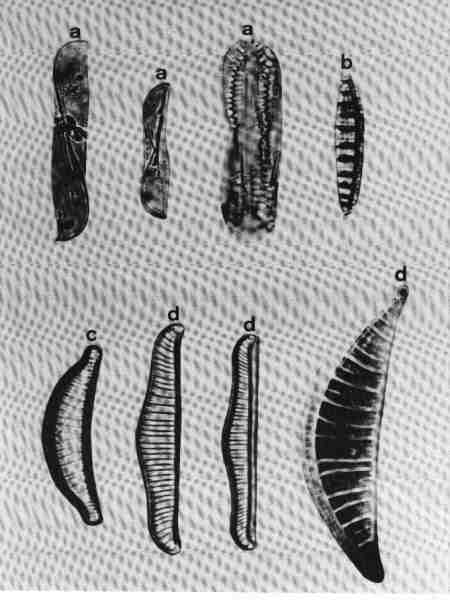
(original size image) |
|
Biraphid Genera
Raphe on sigmoid keel wing
a) Entomoneis - valve view, - girdle view of valve, - girdle view – frustule
Valve crossed by Costae. Eccentric raphe on keel
b) Denticula – valve view
c) Epithemia – valve view
d) Rhopalodia – valve view, - valve view, - valve view (SEM); note the costae across valve
|
c) Raphe on keel wing on valve margin or middle with fibulae
1. Bacillaria (Fig. 21 a)
Cells linked into colonies exhibiting a characteristic 'slide rule' movement in which individual cells slide against each other, through an interlocking ridges and grooves present on the raphe-sternum. Two plastids, one each towards each pole.
Valves linear, linear-lanceolate, with rostrate or capitate poles. Striae parallel consists of single rows of pores separated by ribs. Raphe located almost centrally on Raphe sternum on a keel, supported by arch like fibulae. Raphe extending from pole to pole.
A distinct genus related to Nitzschia, and distributed very commonly in impacted sites of urban streams, epipelic.
2. Hantzschia (Fig. 21 b)
Cells solitary, plastids two to four lying against the ventral side of the whole cell. Valves dorsiventral, or slightly sigmoid, linear-lanceolate with rostrate or apiculate poles. Raphe peripheral located on one margin, both raphes are on the valve margins of the same side of the frustule. Striae made of simple rows of pores parallel, interrupted at the centre, raphe endings simple. Raphe supported by fibulae. This genus can be distinguished from Nitzschia by the location of the raphe on the same side in both valves. The most common species H. ampioxys and H. virgata are universal. It is common in damp soil and streams especially with eroded soil sediments.
3. Nitzschia (Fig. 21 c)
Cells solitary or rarely in chain, plastids two, one on each end of the valve. Valves mostly straight, slightly dorsiventral, rarely sigmoid, narrow, linear, lanceolate or rarely elliptical; with rostrate or capitate ends. Striae mostly parallel consisting of simple areolae to complex loculate structures not interrupted by longitudinal hyaline areas; in most of the species, striae cannot be easily resolved under light microscope. Raphe strongly eccentric peripheral located on one margin. Raphe system on the two valves on opposite margins, axial area very narrow, central area absent. The raphe is on an elevated keel with fibulae.
This is a difficult genus to identify to the species level and is represented by several marine and freshwater species tolerant to pollution, epipelic and benthic. Present in damp soil. Common in streams with sediments from erosion. Research is required to sort out several Australian species.
4. Tryblionella (Fig. 21 d)
Cells solitary, two plastids one on either side of the median transapical axis. Valves linear, elliptical or panduniform with bluntly rounded or apiculate poles. Valve face undulate. Raphe strongly eccentric on a keel on one margin of the valve and a marginal ridge on the other. Striae uniseriate or multiseriate interrupted by one or two hyaline areas. Terminal fissures short and deflected, central external fissures close together slightly expanded.
This genus is very common in the sediments of urban streams of high conductivity and can be distinguished from Nitzschia by its marginal ridges, apically elongate fibulae and longitudinal hyaline areas. Traditionally placed under Nitzschia, this genus has been resurrected by Mann (Round et al.. 1990).
d) Keel raphe on the entire circumference of the valve.
1. Campylodiscus (Fig. 21 e)
Cells curved in girdle view. Valves saddle shaped, subcircular. The raphe system located on keel wing all around the perimeter of the valve and supported by ribs. Striae very fine, intervened by marginal ribs. This is a brackish and freshwater genus easy to identify, represented by one or two species in the urban streams.
2. Stenopterobia (Fig. 21 f)
Valves elongate linear, sigmoid towards the poles, isopolar and isobilateral with rounded ends. Raphe system located on keel wing peripheral around valve margin, similar to Surirella. Striae, fine, with a narrow hyaline area along the apical axis.
This genus is very much like Suirella in structure, but more elongate and linear. This has been found in pristine streams.
3. Surirella (Fig. 21 g)
Cells solitary, one or two plastids – large and highly lobed. Valve linear, elliptic ovate or clavate, isopolar or heteropolar but isobilateral with well rounded, rostrate or apiculate ends. Raphe located on the whole perimeter of the valve on a keel wing supported by rib-like undulations.
Striae fine multiseriate, often separated by transverse ribs extending from the margins, interrupted by 'sterna', (hyaline areas). Raphe simple straight, terminal endings located at the poles. This is common in organic-rich, shallow streams with high conductivity.
| Fig 21 |

(original size image) |
|
Raphe on keel – almost at the middle of valve
Cells Colonial
Bacillaria – valve view. Note the keel at the middle with fibula and keel puncta
Keel on margin – Fibulae and keel puncta
b) Hantzchia
c) Nitzschia – valve view, - girdle view, - valve view
d) Tryblionella – valve view
Keel on the circumference of valve
e) Campylodiscus – valve view
f) Stenopterobia – valve view
g) Surirella – valve view. Fresh cell showing the lobed chloroplastids
|
NOTE
Recent Name Changes
- Species of Sellaphora, Placoneis, Craticula, Fallacia and Hyppodonta were once included under Navicula (Round et al.. 1990, Lange-Bertalot & Metzeltin 1996).
- Several species placed under Achnanthes have been assigned to Achnanthidium (Round et al.. 1990).
- Several species previously assigned to Brachysira have been placed under Anomoeoneis.
- Ctenophora was previously included under Synedra (Round et al.. 1990).
- Several species previously recognised under Cymbella have been included under Encyonema.
- Many species under Fragilaria have been separated and placed under Fragilarioforma, Pseudostaurosira, Staurosira, Staurosirella, Punctastriata (Williams & Round 1987, Round et al.. 1990).
- Tabularia is a genus formerly placed under Synedra, but separated by William & Round 1986.
- The genus Tryblionella was previously placed under Nitzschia (Round et al.. 1990).
- Cylindrotheca is a genus closely related to Nitzschia.
- Nitzschia closterium truly belongs to Cylindrotheca due to its spiral twist (round et al.. 1990).
Chapter 5 Iconographs
Most of the diatom species found in the urban streams are illustrated to show variations. The scale for each micrograph is given so that the user can measure the dimensions and the striae density.
| Fig. 22 Achnanthes/Achnanthidium |

(original size image) |
- Achnanthes brevipes Ag.
Achnanthes brevipes
Achnanthes brevipes
- Achnanthidium binodis
Kütz.
Achnanthidium binodis
Achnanthidium hauckiana Grun.
- Achnanthidium delicatulum Kütz.
Achnanthidium delicatulum
Achnanthidium delicatulum
Scale bar = 10 µm
|
| Fig. 23 Achnanthidium |

(original size image) |
- Achnanthidium minutissima Kütz.
Achnanthidium minutissima
Achnanthidium minutissima (girdle view)
Achnanthidium minutissima (girdle view)
Achnanthidium minutissima (girdle view)
- Achnanthidium oblongella RLV Oestrup
Achnanthidium oblongella RV
- Achnanthidium plonensisRV Hust.
Achnanthidium plonensis RLV
Scale bar = 10 µm
|
| Fig. 24 Amphora |

(original size image) |
- Amphora australiensis John – valve view
Amphora australiensis – frustule (ventral view)
Amphora australiensis
- Amphora coffeaeformis (Ag.) Kütz - valve view
- Amphora fontinalis Hust. – frustule
- Amphora holsatica Hust. – valve view
Amphora holsatica
Amphora holsatica
Scale bar = 10 µm
|
| Fig. 25 Amphora |

(original size image) |
- Amphora ovalis (Kütz.) Kütz – valve view
Amphora ovalis - frustule
Amphora ovalis
Amphora ovalis
Amphora ovalis
Amphora ovalis
- Amphora obtusa Greg.
Scale bar = 10 µm
|
| Fig. 26 Aneumastis/Anomoeneis/Aulacoseira |

(original size image) |
- Aneumastis tusculus (Ehr.) Mann & Stickle
- Anomoeoneis sphaerophora Pfitzer
Anomoeoneis sphaerophora
- Aulacoseira distans (Ehr.) Simonsen
- Aulacoseira granulata (Ehr.) Simonsen
Aulacoseira granulata
Aulacoseira granulata
Scale bar = 10 µm
|
| Fig. 27 Bacillaria/Brachysira |
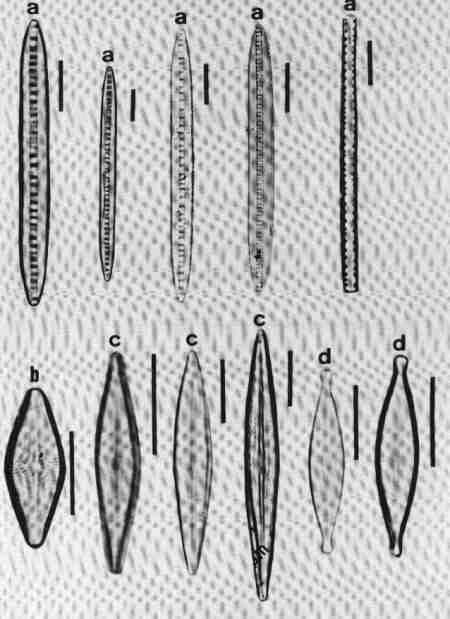
(original size image) |
- Bacillaria paxillifer (O.F Muller) Nitzsch
Bacillaria paxillifer
Bacillaria paxillifer
Bacillaria paxillifer
Bacillaria paxillifer (girdle view)
- Brachysira serians (Breb. ex Kütz.) Round & Mann
- Brachysira styriaca (Grun.) Ross
Brachysira styriaca
Brachysira styriaca
- Brachysira vitrea (Grun.) Ross
Brachysira vitrea
Scale bar = 10 µm
|
| Fig. 28 Caloneis/Cocconeis/Craticula/Ctenophora |
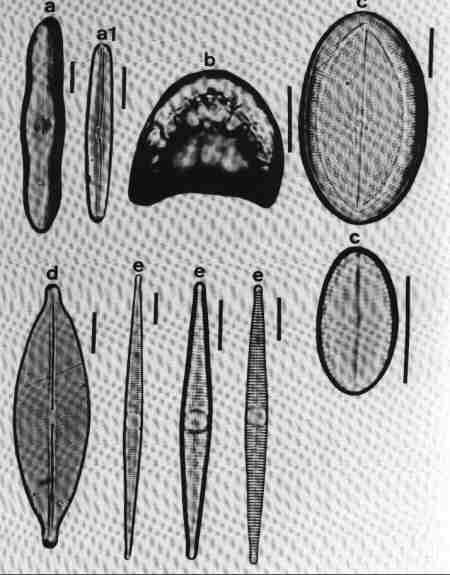
(original size image) |
- Caloneis ventricosa (Ehr.) Meister
a1) Caloneis bacillum
- Campylodiscus clypeus (Ehr.) Ehr.
- Cocconeis placentula RV (Ehr.) Hust.
Cocconeis placentula RLV (Ehr.) Hust.
- Craticula cuspidata (Kütz.) D.G. Mann
- Ctenophora pulchella (Ralfs ex Kütz.) Williams & Round
Ctenophora pulchella
Ctenophora pulchella
Scale bar = 10 µm
|
| Fig. 29 Cyclotella |
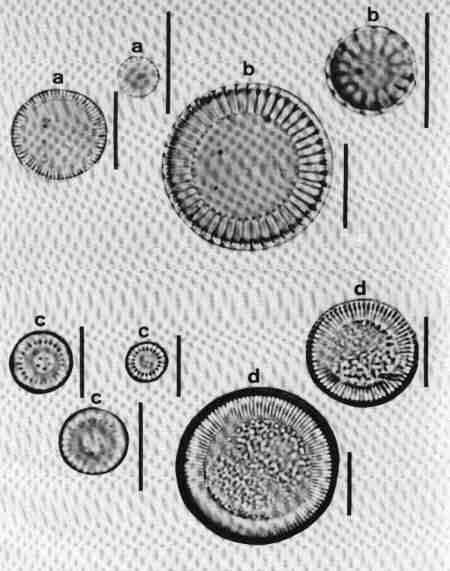
(original size image) |
- Cyclotella atomus Hust.
Cyclotella atomus
- Cyclotella meneghiniana Kütz
Cyclotella meneghiniana
- Cyclotella stelligera Cl. & Grun.
Cyclotella stelligera
Cyclotella stelligera
- Cyclotella striata Kütz & Grun.
Cyclotella striata
Scale bar = 10 µm
|
| Fig. 30 Cymbella |
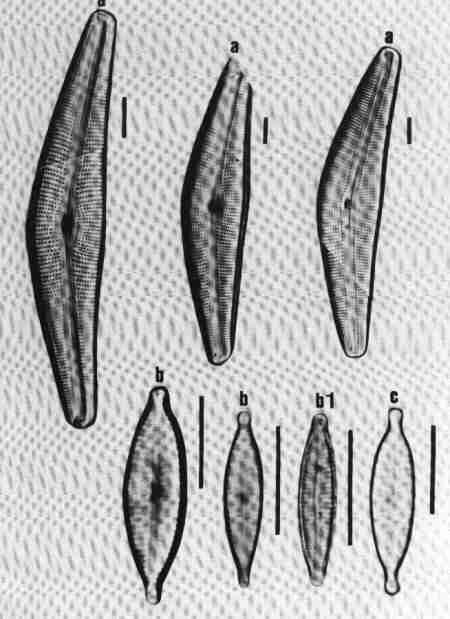
(original size image) |
- Cymbella lanceolata (Ehr.) Van Heurck
Cymbella lanceolata
Cymbella lanceolata
- Cymbella similis Hust.
Cymbella similis
b1) Cymbella microcephala Grun.
- Cymbella similis Hust.
Scale bar = 10 µm
|
| Fig. 31 Cymbella |
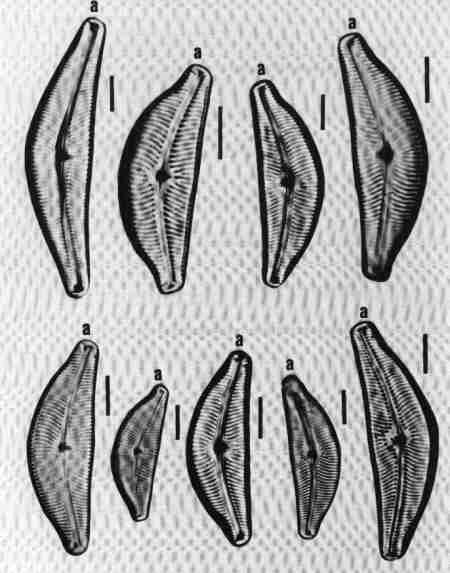
(original size image) |
- Cymbella tumida (Bréb.) Van Heurck
Cymbella tumida
Cymbella tumida
Cymbella tumida
Cymbella tumida
Cymbella tumida
Cymbella tumida
Cymbella tumida
Cymbella tumida
Scale bar = 10 µm
|
| Fig. 32 Denticula/Diadesmis/Diploneis |

(original size image) |
- Denticula subtilis Grun./LI>
- Diadesmis confervacea Kütz
Diadesmis confervacea
Diadesmis confervacea (girdle view)
- Diploneis chersonensis(Grun.) Cl.
- Diploneis ovalis (Hilse) Cl
- Diponeis smithii (Bréb. ex W.Smith) Cl
- Diploneis vacillans (A.Schmidt) Cl
Scale bar = 10 µm
|
| Fig. 33 Encyonema/Entomoneis/Epithemia |
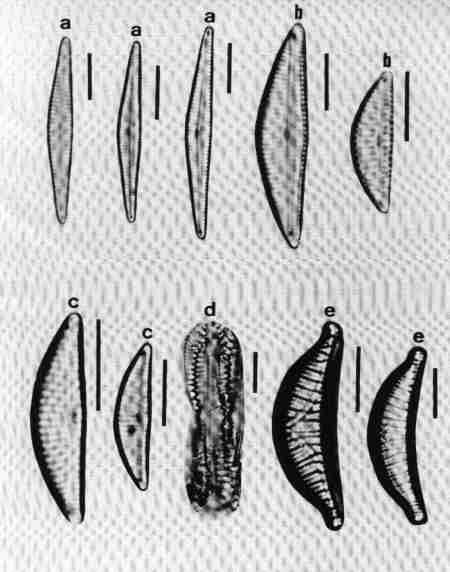
(original size image) |
- Encyonema delicatulum (Kütz.) Mann
Encyonema delicatulum
Encyonema delicatulum
- Encyonema minutum (Hilse) Mann
Encyonema minutum
- Encyonema minutum (Hilse) Mann
Encyonema minutum
- Entomoneis costata
Ehr.
- Epithemia sorex Kütz.
Epithemia sorex
Scale bar = 10 µm
|
| Fig. 34 Eunotia |
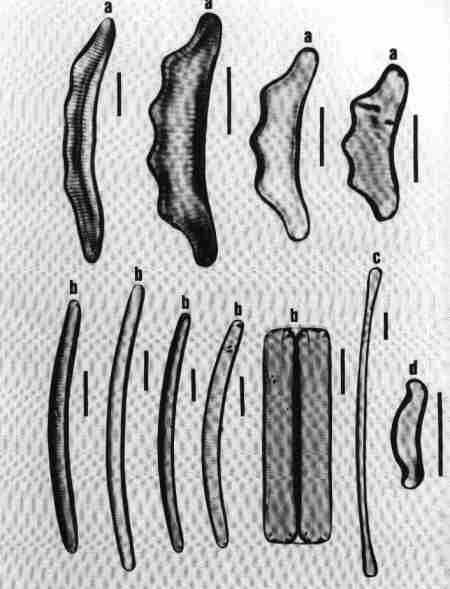
(original size image) |
- Eunotia camelus Ehr.
Eunotia camelus
Eunotia camelus
Eunotia camelus
- Eunotia curvata (Kütz.) Lagerst
Eunotia curvata
Eunotia curvata
Eunotia curvata
Eunotia curvata
Eunotia curvata (girdle view)
- Eunotia eurycephaloides Nörpel-Sch. & Lange-Bertalot
- Eunotia exigua(de Breb.) Grun.
Scale bar = 10 µm
|
| Fig. 35 Eunotia |
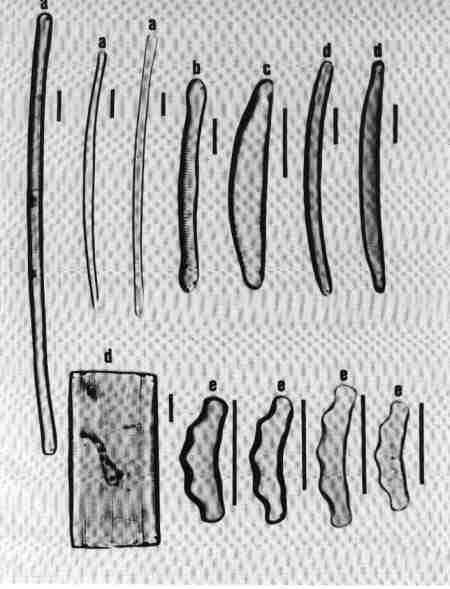
(original size image) |
- Eunotia flexuosa (Breb.) Kütz.
Eunotia flexuosa
Eunotia flexuosa
- Eunotia formica Ehr.
- Eunotia incisa Greg.
- Eunotia pectinalis (Kütz.) Rabh.
Eunotia pectinalis
Eunotia pectinalis (girdle view)
- Eunotia trigibba (Kütz.)
Eunotia trigibba
Eunotia trigibba
Eunotia trigibba
Scale bar = 10 µm
|
| Fig. 36 Fallacia/Fragilaria |

(original size image) |
- Fallacia auriculata (Hust.) Mann
Fallacia auriculata
- Fallacia pygmeae (Kütz.) Stickle & Mann
Fallacia pygmeae
Fallacia pygmeae
- Fragilaria brevistriata
(syn. Pseudostaurosira brevistriata ) (Grun.) William & Round
- Fragilaria punctata John
- Fragilaria rumpens Kütz.
Fragilaria rumpens
e1)Fragilaria vaucheriae (Kütz) Petersen
Fragilaria rumpens
Fragilaria rumpens
Fragilaria rumpens(girdle view)
Fragilaria rumpens (girdle view)
Scale bar = 10 µm
|
| Fig. 37 Frustulia |

(original size image) |
- Frustulia megaliesmontana Cholnoky
Frustulia megaliesmontana
Frustulia megaliesmontana
Frustulia megaliesmontana
Frustulia megaliesmontana
- Frustulia rhomboides
(Ehr.) de Toni
- Frustulia vulgaris (Thwaites) de Toni
Frustulia vulgaris
- Frustulia rhomboides
form capitata
Scale bar = 10 µm
|
| Fig.38 Gomphonema |
(original size image) |
- Gomphonema acuminatum var coronatum (Ehr.) W. Smith
a1) Gomphonema acuminatum var coronatum (Ehr.) W. Smith
Gomphonema acuminatum var coronatum (Ehr.) W. Smith
Gomphonema acuminatum var coronatum (Ehr.) W. Smith
- Gomphonema parvulum (Kütz.) Kütz.
Gomphonema parvulum
Gomphonema parvulum
Gomphonema parvulum
Gomphonema parvulum
Scale bar = 10 µm
|
| Fig.39 Gomphonema |
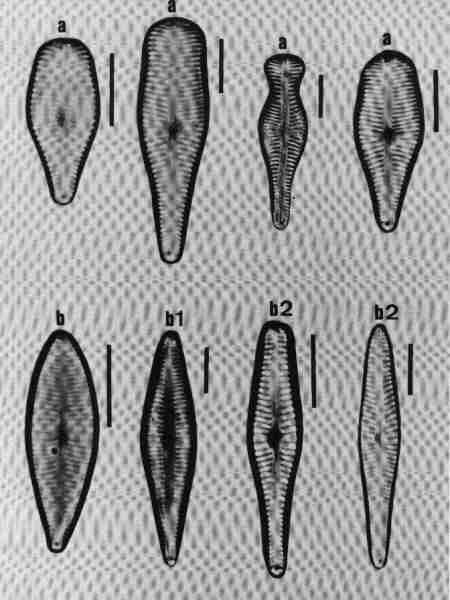
(original size image) |
- Gomphonema truncatum Ehr.
Gomphonema truncatum
Gomphonema truncatum
Gomphonema truncatum
- Gomphonema lanceolatum Ehr.
b1)Gomphonema affine Kütz.
b2)Gomphonema undulatum
- Gomphonema undulatum
Scale bar = 10 µm
|
| Fig.40 Gyrosigma |

(original size image) |
- Gyrosigma spencerii (W. Smith) Cl.
Gyrosigma spencerii
Gyrosigma spencerii
- Gyrosigma balticum (Ehr.) Cl.
- Gyrosigma kutzingii
Gyrosigma kutzingii
Scale bar = 10 µm
|
| Fig.41 Hantzchia/Hyppondonta/Luticola/Martyana |

(original size image) |
- Hantzchia amphioxys (Ehr.) Grun.
- Hantzchia virgata
Hantzchia virgata
Hantzchia virgata
b1) Hantzchia amphioxys (Ehr.) Grun.
- Hyppodonta capitata (Ehr..) Lange-Bertalot
Hyppodonta capitata
- Luticola mutica (Kütz) Mann
- Luticola pseudomutica (Hust.) Mann
- Martyana martyi (Heribaud) Round
Scale bar = 10 µm
|
| Fig.42 Mastogloia |

(original size image) |
- Mastogloia braunii Grun.
Mastogloia braunii
Mastogloia braunii
Mastogloia braunii
Mastogloia braunii
Mastogloia braunii
- Mastogloia elliptica (Ag.) Cl.
- Mastogloia smithii Thwaites
Mastogloia smithii
Scale bar = 10 µm
|
| Fig.43 Melosira/Meridion |
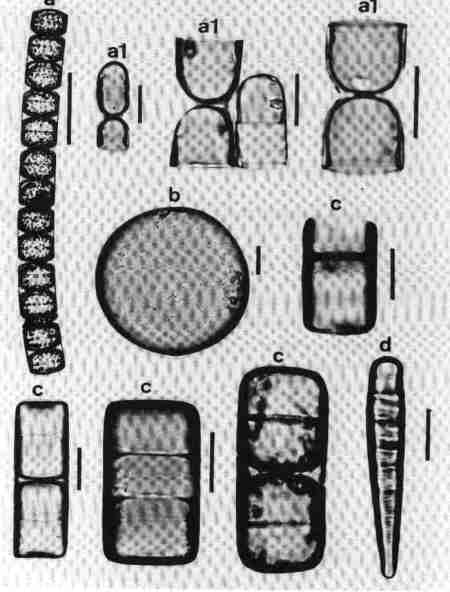
(original size image) |
- Melosira moniliformis (Müller) Ag. Fresh cells in chain.
Scale = 100 µm
a1) Melosira nummuloides (Dillwyn) Ag.
Melosira nummuloides
Melosira nummuloides
- Melosira moniliformis (Müller) Ag. - valve view
- Melosira varians Ag.
Melosira varians
Melosira varians
Melosira varians
Melosira varians
- Meridion circulare (Grev.) Ag
Scale bar = 10 µm
|
| Fig.44 Navicula |

(original size image) |
- Navicula accomoda Hust.
Navicula accomoda
- Navicula cancellata Donkin
- Navicula cincta (Ehr.) Kütz
Navicula cincta
Navicula cincta
Navicula cryptocephala Kütz
Navicula cryptocephala
Navicula cryptocephala
Navicula cryptocephala
Navicula cryptocephala
- Navicula elginensis
(Greg.) Grun
Scale bar = 10 µm
|
| Fig.45 Navicula |
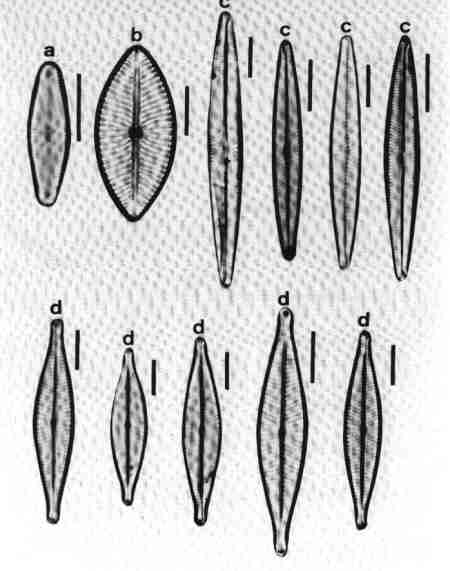
(original size image) |
- Navicula festiva Krasske
- Navicula gastrum (Ehr.) Donkin
(syn Placoneis gastrum) (Ehr.) Mereschkowsky
- Navicula radiosa Kütz.
Navicula radiosa
Navicula radiosa
Navicula radiosa
- Navicula rhynchocephala Kütz.
Navicula rhynchocephala
Navicula rhynchocephala
Navicula rhynchocephala
Navicula rhynchocephala
Scale bar = 10 µm
|
| Fig.46 Navicula |

(original size image) |
- Navicula viridula Kütz
Navicula viridula
- Navicula salinarum Grun.
- Navicula stigmatifera John
- Navicula rhynchocephala
- Navicula symmetrica Patrick
- Navicula tripuncata Kütz
Navicula tripuncata
Navicula tripuncata
Navicula tripuncata
- Navicula viridula Kütz
Scale bar = 10 µm
|
| Fig.47 Neidium |
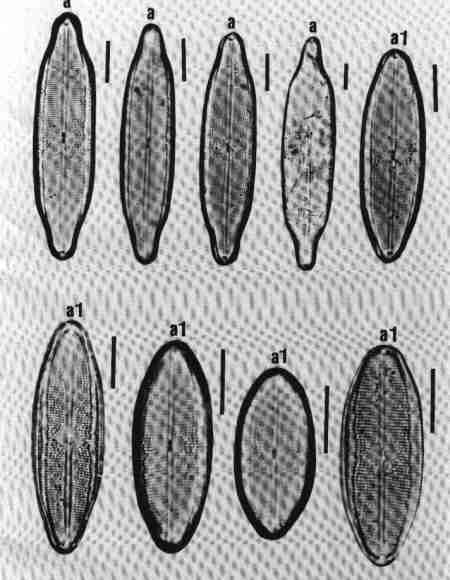
(original size image) |
a) Neidium productum (W. Smith) Cl.
Neidium productum
Neidium productum
Neidium productum
a1) Neidium affine Ehr. Pfitz
Neidium affine
Neidium affine
Neidium affine
Neidium affine
Scale bar = 10 µm
|
| Fig.48 Nitzschia |
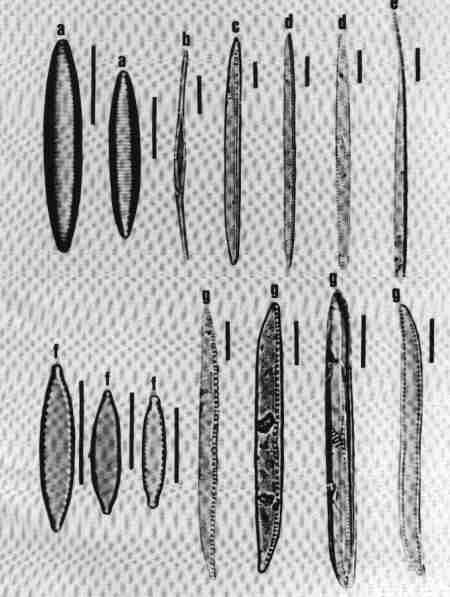
(original size image) |
- Nitzschia amphibia Grun.
Nitzschia amphibia
- Nitzschia closterium (Ehr). W. Smith
(Syn. Cylindrotheca closterium)
- Nitzschia dissipata (Kütz) Grun.
- Nitzschia linearis W. Smith
Nitzschia linearis
- Nitzschia lorenziana Grun.
- Nitzschia microcephala Grun.
Nitzschia microcephala
Nitzschia microcephala
- Nitzschia obtusa W. Smith
Nitzschia obtusa
Nitzschia obtusa
Nitzschia obtusa
Scale bar = 10 µm
|
| Fig.49 Nitzschia |
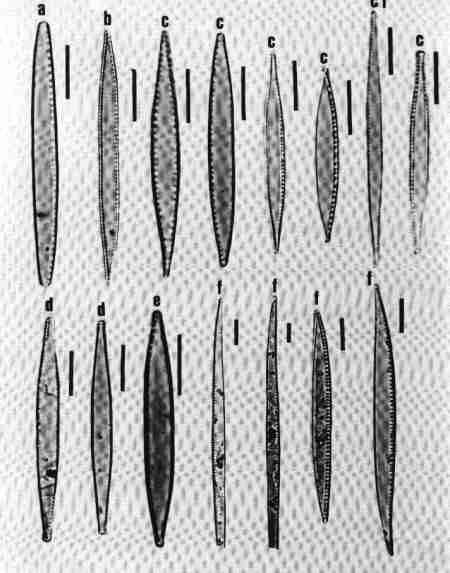
(original size image) |
- Nitzschia obtusa
- Nitzschia gracilis Hantzch
- Nitzschia palea (Kütz.) W. Smith.
Nitzschia palea
Nitzschia palea
Nitzschia palea
c1.Nitzschia acicularis
Nitzschia palea
- Nitzschia sp.
Nitzschia sp.
- Nitzschia paleacea Grun.
- Nitzschia sigma (Kütz) W. Smith
Nitzschia sigma
Nitzschia sigma
Nitzschia sigma
Scale bar = 10 µm
|
| Fig.50 Paralia/Pinnularia |

(original size image) |
- Paralia sulcata (W. Smith) Crawford
- Pinnularia borealis Ehr.
- Pinnularia subcapitata Greg.
- Pinnularia maior Ehr.
- Pinnularia divergens W. Smith
- Pinnularia gibba Ehr.
Pinnularia gibba
Pinnularia gibba (girdle view)
- Pinnularia microstauron (Ehr.) Cl.
- Pinnularia viridis (Nitz.) Ehr.
Scale bar = 10 µm
|
| Fig.51 Pleurosigma |

(original size image) |
- Pleurosigma elongatum W. Smith
- Pleurosigma longum Perg. Scale = 50 µm
b1) Pleurosigma longum
- Pleurosigma salinarum Grun. Scale = 50 µm
Pleurosigma salinarum
Scale bar = 10 µm
|
| Fig.52 Rhopalodia |
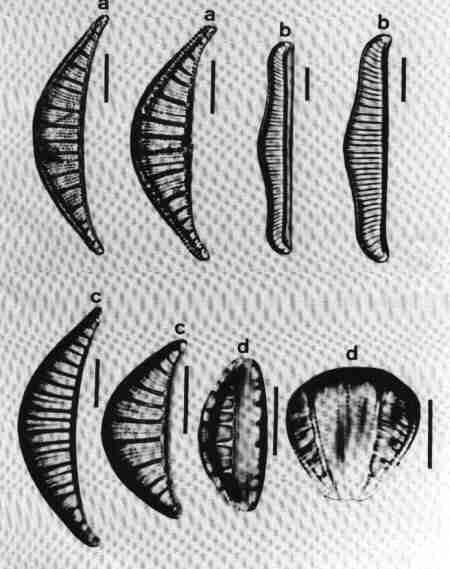
(original size image) |
- Rhopalodia brebissoni Krammer
Rhopalodia brebissoni
- Rhopalodia gibba (Ehr.) O. Müller
Rhopalodia gibba
- Rhopalodia gibberula
(Ehr.) O. Müller
Rhopalodia gibberula
- Rhopalodia musculus
(Kütz ) O. Müller
Rhopalodia musculus - Frustule
Scale bar = 10 µm
|
| Fig.53 Sellaphora |
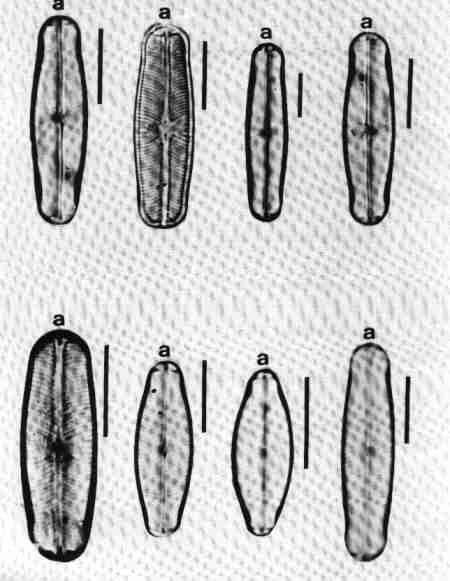
(original size image) |
- Sellaphora pupila (Kütz.) Mereschkowsky
Sellaphora pupila
Sellaphora pupila
Sellaphora pupila
Sellaphora pupila
Sellaphora pupila
Sellaphora pupila
Sellaphora pupila
Scale bar = 10 µm
|
| Fig.54 Skeletonema/Stauroneis/Stenopterobia |
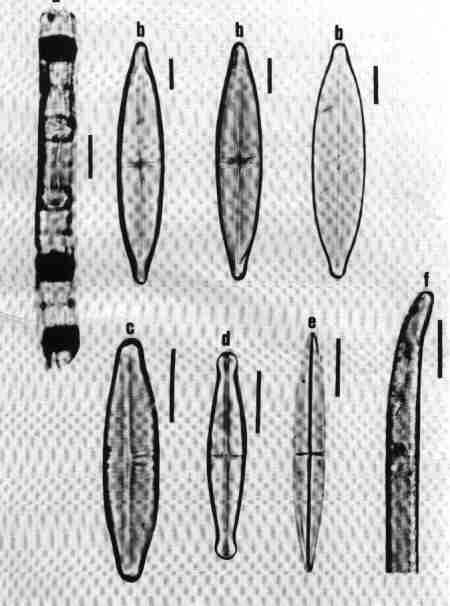
(original size image) |
- Skeletonema costatum (Grev.) Cl.
- Stauroneis anceps Ehr.
Stauroneis anceps
Stauroneis anceps
- Stauroneis dubitabilis Hust.
- Stauroneis pachycephala Cl.
- Stauroneis spicula Hickie
- Stenopterobia sigmatella (Gregory) Ross
Scale bar = 10 µm
|
| Fig.55 Staurosira/ Surirella |

(original size image) |
- Staurosira construens(Ehr.) Williams and Round
- Surirella angusta Kütz
Surirella angusta
Surirella angusta
Surirella angusta
Surirella angusta
- Surirella ovalis Bréb
Surirella ovalis
- Surirella tenera Greg.
Scale = 10 µm
|
| Fig. 56 Synedra/Tabularia |

(original size image) |
- Synedra ulna (Nitsch) Ehr.
Synedra ulna
Synedra ulna
Synedra ulna
Synedra ulna
Synedra ulna
Synedra ulna
Synedra ulna
Synedra ulna
- Tabularia tabulata (Ag.) Snoeijs
(Syn. T. fasciculata) (Ag) Williams & Round
Tabularia tabulata
Scale = 10 µm
|
| Fig. 57 Tabellaria |
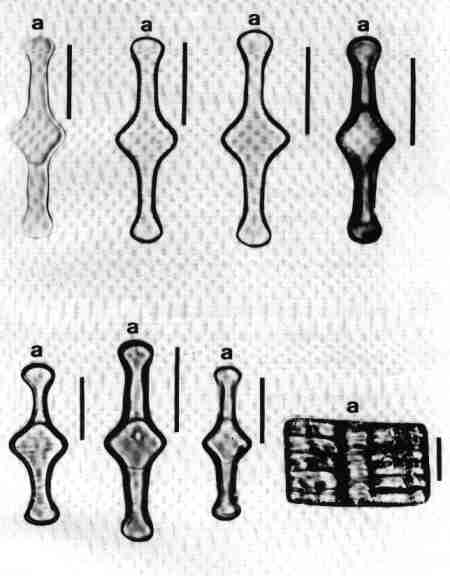
(original size image) |
- Tabellaria flocculosa (Rabh.) Kütz
Tabellaria flocculosa
Tabellaria flocculosa
Tabellaria flocculosa
Tabellaria flocculosa
Tabellaria flocculosa
Tabellaria flocculosa
Tabellaria flocculosa (girdle view)
Scale = 10 µm
|
| Fig. 58 Thalassionema |
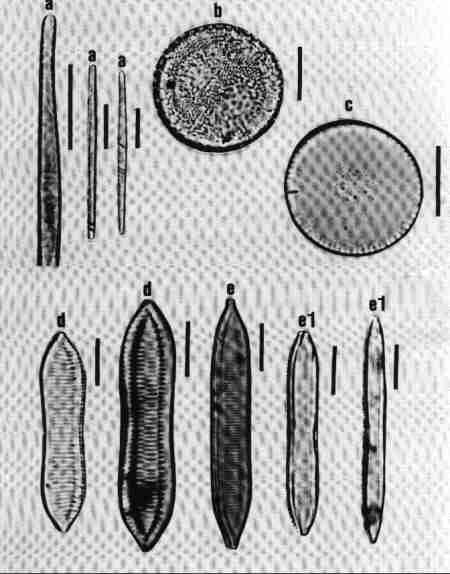
(original size image) |
- Thalassionema nitzschiodes Grun.
Thalassionema nitzschiodes
Thalassionema nitzschiodes
- Thalassiosira australis Grun.
(Syn. Thalassiosira lacustris) (Grun.) Hasle
- Thalassiosira weissflogii (Grun.) Fryxell & Hasle
- Tryblionella levidensis (W. Smith) Heurck
(Syn. Nitzschia tryblionella var levidensis) (W. Smith) Grun.
Tryblionella levidensis
- Tryblionella levidensis
e1) Tryblionella apiculata
Tryblionella apiculata
Scale = 10 µm
|
| Fig. 59 Tryblionella |

(original size image) |
- Tryblionella levidensis (W. Smith) Heurck
- Tryblionella hungarica Grun.
- Tryblionella circumsuta (Bail) Grun.
- Tryblionella victoriae Grun.
(Syn Nitzschia tryblionella var victoriae) Grun.
Tryblionella victoriae
Tryblionella victoriae
Tryblionella victoriae
Scale = 10 µm
|
References
- Anderson, W.J. (1993). Natural versus anthropogenic change in lakes: the role of sediment record. Trends in Ecology and Evolution 8: 356-361
- Archibald,R.E.M. (1983). The diatoms of the Sundays and Great Fish rivers in the eastern cape province of South Africa. Bibliotheca diatomologica. J. Cramer. Stuttgart 361pp.
- Bateman, L. and Rushforth, S.R. (1982). Bibliotheca diatomologica 4. J. Cramer, Stuttgart. 99pp.
- Blaze, K.L. (1984). Morphology and taxonomy of Diplomenora gen:nov. (Bacillariophyta). Br. Phycol. Journal. 19: 217-225
- Blinn, D.W. (1995). Diatom community structure along salinity gradients in Australian saline lakes: biogeographic comparisons with other continents. In: Kociolek, J.P. and Sullivan, M.J. (eds) A Century of Diatom Research in North America: A Tribute to the Distinguished Careers of Charles W. Reimer and Ruth Patrick. Koeltz Scientific Books, USA. pp 163-174.
- Camburn, K.E., Kingston, J.C. and Charles, D.F. (1984-86). Pirla unpublished report series 3. Pirla, Diatom Iconograph.
- Cassie, V. (1989). A contribution to the study of New Zealand diatoms. Bibliotheca diatomologica 17. J. Cramer, Stuttgart. 266pp.
- Chessman, B., Gowns, I., Currey, J. and Plunkett-Cole, N. (1999). Predicting diatom communities at the genus level for rapid biological assessment of rivers. Freshwater Biology 41(2): 317-333.
- Christie, C.E. and Smol, J.P. (1993). Diatom assemblages as indicators of lake trophic status in the south-eastern Ontario lakes. Journal of Phycology 29: 575-586.
- Clark, R.L. and Rushforth, S.R. (1977). Diatom studies of the headwaters of Henry's fork of the Snake river Island Park. Idaho USA. Bibliotheca phycologia 33. J Cramer. 204pp.
- Cox. E.J. (1996). Identification of freshwater diatoms from live material. Chapman Hall. 158pp.
- Czarnecki, D.B. and Blinn, D.W. (1978). Diatoms of the Colorado River in Grand Canyon National Park and vicinity. Bibliotheca phycologia 38. J Cramer, Vaduz. 181pp.
- Day, S.A., Wickham, R.P., Entwisle, T.J. & Tyler, P.A. (1995). Bibliographic Checklist of Non-marine Algae in Australia. Flora of Australia, Supplementary Series (4). 276pp.
- Dodd, J.H. (1987). The illustrated flora of Illinois Diatoms. Southern Illinois University Press, Carbondale. 477pp.
- Domitrovic, Y.Z.D. and Maidana, N.I. (1997). Taxonomic and ecological studies of the Panama river diatom flora, Argentina. ). Bibliotheca diatomologica 34. J. Cramer, Stuttgart. 122pp.
- Edgar & Pickett-Heaps, J.D. (1983). The mechanism of diatom locomotion: 1. An ultrrstructural study of the motility apparatus. Prodeedings of the Royal Society of London. B. 281. 331-343.
- Edlund, M.B. & Stoermer, E.F. (1997). Ecological evolutionary and systematic significance of diatom life histories. Journal of Phycology. 33, 897-918.
- Foged, N. (1976). Freshwater diatoms in Sri Lanka. Bibliotheca phycologia. J Cramer, Vaduz. 113pp.
- Foged, N. (1978). Diatoms in Eastern Australia: Bibliothea phycologia. 41. J. Cramer., Vaduz. 243pp.
- Foged, N. (1984). Freshwater and littoral diatoms from Cuba. Bibliotheca diatomologica 5. J. Cramer, Vaduz. 243pp.
- Foged, N. (1987). Diatoms fron Viti Levu, Fiji Islands. Bibliotheca diatomologica 14.
- J. Cramer, Vaduz.
- Frenguelli, J. (1923- 1924). Diatomeas Argentinas. Reprint 1976, Linnaeus Press. Amsterdam.
- Fritz, S.C., Juggins, S. and Battarbee, R.W. (1993). Diatom assemblages and ionic characterisation of lakes of the Northern great plains, North America. A tool for reconstructing past salinity and climate fluctuations. Canadian Journal of Fisheries and Aquatic Science 50; 1844-1856.
- Fritz, S.C., Cumming, B.F., Gasse, F. and Laird, K.R. (1999). Diatoms as indicators of hydrologic and climatic change in saline lakes. In: Stoermer, E.F. and Smol, J.P. (ed) The Diatoms: Applications for the Environmental and Earth Sciences. Cambridge University Press, England. pp. 41-72.
- Fryxell, G.A. and Villac, M.C. (1999). Toxic and harmful marine diatoms. In: Stoermer, E.F. and Smol, J.P. (ed) The Diatoms: Applications for the Environmental and Earth Sciences. Cambridge University Press, England. pp. 419-428.
- Gasse, F. (1986). East African Diatoms. Taxonomy, ecological distributions. Bibliotheca diatomologica II. J.Cramer., Stuttgart. 313pp.
- Gell, P.A. (1997). The development of a diatom database for inferring lake salinity towards a quantitative approach for reconstructing past climates. Australian Journal of Botany 45: 389-423.
- Gell, P.A. (1998). Quantitative reconstructions of the Holocene palaeosalinity of paired crater lakes based on a diatom transfer function. Palaeoclimates. 31(1-3). 83-96.
- Germain, H. (1981). Flore des diatomees diatomophycees societe nouvelle des editions Boubee. Paris. 441pp.
- Hallegraeff, G.M. (1992). Observations on the mucilaginous diatom Thalassiosira bipartita (Rattray) comb. nov. from the tropical Indo-West Pacific. Diatom Research. 7, No. I 15-23.
- Hall, R.I. and Smol, J.P. (1999). Diatoms as indicators of lake eutrophication. In: Stoermer, E.F. and Smol, J.P. (ed) The Diatoms: Applications for the Environmental and Earth Sciences. Cambridge University Press, England. pp. 128-168.
- Hansman, E.W. (1973). Diatoms of the streams of eastern Conneticut. Dept of Environmental Protection, Bulletin 106. 119pp.
- Harding, J.P.C. & Kelly, M.G. (1999). Recent developments in algal based monitoring in the United Kingdom. In: Prygiel, J., Whitton, B.A. & Bukowska, J. (eds) Use of algae for monitoring rivers III. Agence de l'eau Artois Picardie, France. pp 26-34
- Harwood, D.M. (1999). Diatomite. In: Stoermer, E.F. and Smol, J.P. (ed) The Diatoms: Applications for the Environmental and Earth Sciences. Cambridge University Press, England. pp. 436-446.
- Hayworth, E.Y. & Tyler, P.A. (1993). Morphology and taxonomy of Cyclotella tasmanica sp. nov a newly described diatom from Tasmanian lake. In: Van Dam (ed) 12th International Diatom Symposium. Hydrobiologia 269/270: 47-56.
- Hodgson, D., Vyverman, W. & Tyler, P. (1997). Diatoms of meromictic lakes adjacent to the Gordon River and of the Gordon River estuary in south-west Tasmania. Bibliotheca diatomologica. 35. J. Cramer, Stuttgart. 173pp.
- Holland, J. & Clark, R.L. (1989). Diatoms of Burrinjuck Reservoir, New South Wales, Australia. CSIRO Divisional Report 89/1. Institute of Natural Resources and Environment. Division of Water Resources. 71 pp.
- Hustedt, F. (1927-1966). Die Kieselalgen Deutschlands, Österrerchs und der Schweiz mit Berücksichtigung der übrigen lander Europas sowie der angrenzenden Meeresgebiete. In: Rabenhorst, L. Kryptogamen-Flora, Band VII. Teil, 1,2& 3. Reprint 1977.
- Hustedt, F. (1930). Bacillariophyta In: Pascher, Süßwasserflora von Mitteleuropa, Heft 10: 1-466. Jena.
- John, J. (1980). Two new species of diatom, Mastogloia from Western Australia. Nova Hedwigia 33: 849-858. Braunschweig, Germany.
- John, J. (1981a). New species of freshwater diatoms from Western Australia. . Nova Hedwigia 34: 569-576. Germany.
- John, J. (1981b). Amphora australiensis sp. nov. Nova Hedwigia 35: 39-53. Germany.
- John, J. (1981c). Three new taxa of diatoms from Western Australia. Cyptogamie Algolgie 11 (2): 131-139. Paris.
- John, J. (1982a). Valve structure of Cymbella sumatrensis Hust. From Northern Australia. Bacillaria 5: 117-126. Bremerhaven, Germany.
- John, J. (1982b). Observations on Hyalodiscus lentiginous sp. nov. from Western Australia. Nova Hedwigia 37: 709-721. Germany.
- John, J. (1983a). The Diatom Flora of the Swan River Estuary, Western Australia, Bibliotheca Phycologia. 64: 1-359. J. Cramer., Veduz. 360pp.
- John, J. (1983b). Observations on Thalassiosira lacustris (Grun.) Hasle. Nova Hedwigia 38: 337. Germany.
- John, J. (1986). Observations on the ultra-structure of Gephyria media W. Arnott. In: Ricard, M. (ed) Proceedings of the 8th International Diatom Symposium. 155-162, Koetz Scientific Books, Koenigstein, Germany.
- John, J. (1987). (ed). Swan River estuary, ecology and management. Environmental Studies Group Report I, Curtin University of Technology.
- John, J. (1988). Observations on the morphology and ecology of Navicula elegans from Western Australia. In: Proceedings of the 9th International Diatom Symposium. F.E. Round: (ed.) 406-417, Bio Press Ltd, UK.
- John, J. (1989). Distributions of epiphytic diatom communities in the Swan River estuary in relation to hydrological factors. In: Proceeding of the10th International Diatom Symposium. H. Simola. (ed.), Finland.
- John, J. (1990a). Diatoms associated with stromatolites at Shark Bay, Western Australia. In:Special vol. Dedicated to Dr Germain. (ed.). M Ricard. Nat. Hist. Museum. Paris.
- John, J. (1990b). Parlibellus panduriformis, a new species of diatom from Shark Bay, Western Australia. Phycologia (30)6: 38-42.
- John, J. (1990c). An epizoic diatom from the Swan River estuary. Journal of the Royal Society of Western Australia. 73(3): 73-75
- John, J. (1991). Climaconeis stromatolitis, a new species of diatom from Shark Bay, Western Australia. Diatom Research 6(1): 49-54.
- John, J. and Economou-Amilli,A. (1991). Morphology and ultrastructure of the centric diatom Cyclotella distinguenda. Diatom Research 6(2): 307-315.
- John, J. and van Riessen. A. (1992). Mucilage structures of Mastogloia fimbriata and Mastogloia cocconeiformis examined by the environmental scanning electron microscope. Diatom Research 7(2): 403-407.
- John, J. (1993a). Morphology and ultrastructure of the tubes of Mastogloia cocconeiformis (Bacillariophyta) from Shark Bay, Western Australia. Phycologia 32(5).
- John, J. (1993b). The use of diatoms in monitoring the development of created wetlands at RGC mine site, Capel, Western Australia. Hydrobiologia 269/270; 427-436.
- John, J. (1993c). Palaeolimnology of North Lake, Perth, Western Australia, based on diatom assemblages. In: Bowdrey, G. and Hope, G.S.(ed) Sixth International Palaeolimnology Symposium, Australia National University, Canberra. pp49
- John, J. (1994). Eutrophication of the Swan River and Management Strategy. In: Global Wetlands. Elsevier, New York.
- John, J. (1998). Diatoms: Tools for Bioassessment of River Health. Report to Land and Water Resources Research and Development Corporation, Canberra. 388pp
- John, J. (2000). Diatom prediction and classification system for urban streams. LWRRDC Canberra.
- Krammer, K. and Lange Bertalot, H. (1986). Bacillariophyceae. 1. Teil: Naviculaceae. Band 2/1 Von: Susswasse flora von Mitteleuropa Ettl, H., Gerloff, J., Heynig, H. and Mollenhauer, D., Fisher., Jena (eds). 876pp.
- Krammer, K. and Lange Bertalot, H. (1989). Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiaceae, Suirellaceae, Band 2/2 Susswasser flora von Mitteleuropa (eds) ., Ettl, H Gerloff, J., Heynig, H. and Mellenhaner, D., Fisher., Stuttgart and Jena. 583pp.
- Krammer, K. and Lange Bertalot, H.(1991). Bacillariophyceae. 3. Teil: Centrales, Fragilariaceae, Eunotiaceae, Band 2/3 Von: susswassr flora von Mitteloruopa (eds) ., Ettl, H Gerloff, J., Heynig, H. and Mollenhauer, D., Fisher., Stuttgart and Jena. 576pp.
- Lange-Bertalot, H & Krammer (1989). Achnanthes. Bibliotheca diatomologica. 18. Stuttgart. 393pp.
- Lange-Bertalot,H., Kulbst, K., Lanser, T., Norpelm Schempp, M. and Willmann, M. (1996). Diatom data introduced by Georg Krasske, documentation and revision. Iconographia diatomologica, 3. Koeltz Scientific Books. 358pp.
- Lange-Bertalot, H. and Metzeltin, D. (1996). Indicators of Oligotrophy. 800 taxa representative of three ecological distinct lake types – carbonate buffered, oligodysrophic, weakly buffered soft water. Iconographia Diatomologica 2. Koeltz Scientific Books, Konigstein. 390pp.
- Lange-Bertalot, H. (1979) Pollution tolerance of diatoms as a criterion for water quality estimation. Beih Nova Hedwigia 64: 285-304.
- Lange-Bertalot, H. (1996). Iconographia Diatomologica 4. Koeltz Scientific Books, Konigstein. 286pp.
- Lee, JJ. and Xenophontes, X. (1990). The unusual lifecycle of Navicula muscatinei. Diatom Research 4: 69-77.
- Lokman, S. (1998). Biochemical composition of the local diatom Skeletonema isolate used in Malaysian aquaculture as live food. In: John, J. (ed) 15th International diatom symposium abstracts. pp59.
- Mann, D.G. (1989). The diatom genus Sellaphora separate from Navicula. Br. Phycological Journal 24: 1-20.
- Mann, D.G. (1999). The species concept in diatoms. Phycologia 38 (6), 437-495.
- McBride, P. and Selkirk, J (1998). Palaeo lake diatoms on subarntartic Macquarie Island: possible markers of climate change. In: John, J. (ed) 15th International diatom symposium abstracts.
- Metzeltin, D. and Lange-Bertalot, H. (1998). Tropical diatoms of South America Iconographia Diatomologica 5, Koeltz Scientific Books, 695 pp.
- Metzeltin, D. and Wilkowski, A. (1996). Diatomeen der Baren-Insel. Iconographia diatomologica 4. Koeltz Scientific Books, Konigstein. 286pp.
- Patrick, R. & Reimer, C.W. (1966). The diatoms of the United States Vol. 1. Monographs of the academy of natural sciences of Philadelphia. 13. 688pp.
- Patrick, R. & Reimer, C.W. (1975). The diatoms of the United States Vol. 2. Monographs of the academy of natural sciences of Philadelphia. 13. 213pp.
- Peabody, A.J. (1971). A case of safe breaking involving diatomaceous safe ballast. Journal of Forensic Science Society 11: 227
- Peabody, A.J. (1977). Diatoms in forensic science. Journal of Forensic Science Society. 17: 81-87.
- Peabody, A.J. (1980). Diatoms and drowning- a review – Forensic Science 1: 397-409.
- Peabody, A.J. (1999). Forensic science and diatoms. . In: Stoermer, E.F. and Smol, J.P. (ed) The Diatoms: Applications for the Environmental and Earth Sciences. Cambridge University Press, England. pp. 413-418.
- Pickett-Heaps, J.D., Tippit, D.H. and Leslie, R. (1980). Light and electron microscopic observations on cell division in two large pennate diatoms, Hantzchia and Nitzschia. Mitosis in vivo. European Journal of Cell Biology. 21: 1-11.
- Pickett-Heaps, J.D. (1987). Diatom mitosis implications of a model system. In: Wiessner, W., Robinson, D.G. and Starr, R.C. (eds) Algal Development. Springer –Verlag, Berlin. pp 28-33.
- Podozorski, A.C. and Hakansson, H. (1987). Freshwater and marine diatoms from Palawan. Bibliotheca diatomologica. 13. J. Cramer, Stuttgart. pp245.
- Podzorski, A.C. (1983). The reaction of epiphytic diatoms (Bacillariophyta) to environmental change in Broad River, Western Jamaica. Nova Hedwigia 40: 487-509.
- Prygiel, J. and Coste (1999). Progress in the use of diatoms for monitoring rivers in France. In: Prygiel, J., Whitton, B.A. and Bukoswska (eds) Use of algae for monitoring rivers III. pp 165 – 181.
- Reichardt, E. (1984). Die Diatomeen der almuhe. Bibliotheca diatomologica, J. Cramer. Vaduz. 69pp.
- Round, F.E., Crawford, R.M. and Mann, D.G. (1990). The Diatoms. Cambridge University Press, New York. 747pp.
- Schmid, A.M.M. (1994). Aspects of morphogenesis and function of diatom cell walls with implications for taxonomy. Protoplasma 181: 43-50.
- Schmidt et al (1874-1959). Atlas der diatomaceen-kunde. Band 1, II & III. Tafel 1- 480. Reprint 1984. Koeltz scientific books Koenigstein. Schmidt, A. Schimdt, M. Fricke, F., Heiden, H., Muller, O and Hustedt, F.
- Schoeman, F.R. and Archibald, F.R. (1976 – 1979). The diatom flora of Southern Africa, National Institute for Water Research.
- Schoeman, F.R. (1973). A systematic and ecological study of the diatom flora of Lesotho with special reference to water quality, V.R. Printers, Pretoria pp.355.
- Sims (ed). (1996). An atlas of british diatoms arranged by Hartley, B. Biopress Ltd. 601pp.
- Snoeijis, P. and Vilbaste, S. (1994). Intercalibration and distribution of diatom species in the Baltic sea 2. Opulus Press, Uppsala. 126pp.
- Snoeijis, P. (1993) ). Intercalibration and distribution of diatom species in the Baltic sea. Vol 1. Baltic marine biologists publication 16a. Opulus Press, Lippsala. 129pp.
- Snoeijs, P. (1999). Diatoms and environmental change in brackish waters. In: Stoermer, E.F. and Smol, J.P. (ed) The Diatoms: Applications for the Environmental and Earth Sciences. Cambridge University Press, England. pp. 299-333.
- Stevenson, R.J. and Pan, Y. (1999). Assessing environmental conditions in rivers and streams with diatoms. In: Stoermer, E.F. and Smol, J.P. (ed) The Diatoms: Applications for the Environmental and Earth Sciences. Cambridge University Press, England. pp. 11-40.
- Sullivan,M.J. (1999). Applied diatom studies in estuaries and shallow coastal environments. In: Stoermer, E.F. and Smol, J.P. (ed) The Diatoms: Applications for the Environmental and Earth Sciences. Cambridge University Press, England. pp. 334-351.
- Thomas,D.P. (1983). A limnological survey of Alligators Rivers Region, Northern Territory. Part I Diatoms. Australian Government Publishing Service. 140pp.
- Stosch, H.A. von (1975). An amended terminology of diatom girdle. Nova hedwigia. Beih. 53: 1 – 35.
- Vyverman, W., Vyverman, R., Hodgson, D. & Tyler, P. (1995). Diatoms from Tasmanian mountain lakes: a reference data set (TAS DIAT) for environmental reconstruction and a systematic autecological study. Bibliotheca. Diatomologica. 33. 193pp.
- Vyverman, W., Sabbe, K. & Vyverman, R. (1997). Five new freshwater species of Biremis (Bacillariophyta) from Tasmania. Phycologia. 36: 91-102.
- Vyverman, W., Sabbe, K., Mann, D.G., Kilroy, C., Vyverman, R., Van houtte, K. & Hodgson, D. (1998). Eunophora gen. nov (Bacillariophyta) from Tasmania and New Zealand: description and comparison with Eunotia and amphoroid diatoms. European Journal of Phycology, 33: 95-111.
- Wilkowski (1994). Recent and fossil diatoms flora of the Gulf of Gdansk, Southern Baltic Sea. Bibliotheca diatomologica. 28. J. Cramer, Stuttgart. 313pp.
- Williams, D.M and Round, F.E. (1987). Revision of the genus Fragilaria. Diatom Research, 2: 267-288.
- Wood, E.J.F., Crosby, L.H. and Cassie, V. (1959). Studies on Australian and New Zealand diatoms III. Descriptions of further discoid species. Trans. Roy. Soci. New Zealand 87 (3,4): 211 – 219.
- Wood, E.J.F. (1961a ). Studies on Australian and New Zealand diatoms IV. Descriptions of further sedentary species. Trans. Roy. Soci. New Zealand 33(4): 699-698.
- Wood, E.J.F. (1961b). Studies on Australian and New Zealand diatoms V. The Rawson collection of recent diatoms. Trans. Roy. Soci. New Zealand 88(4): 699-712.
- Wood, E.J.F. (1963a.) Checklist of diatoms recorded from the Indian Ocean. CSIRO, Division of Fisheries and Oceanography. Report 36. Marine Laboratory , Cronulla, Sydney. 304pp.
- Wood, E.J.F. (1963b). Checklist of diatoms recorded from the Indian Ocean. Report 36. C.S.I.R.O., Marine Laboratory, Cronulla, Sydney: 1-304.
- Wood, E.J.F. (1964). Ecological Relations of Australian Estuarine Diatoms. Nov. Hedwigia 8(3/4): 527-548.
Diatom Index
An Index of diatoms appearing in this document (the hard copy has matching page numbers - B Hall)
|
Achnanthes
- A.brevipes
- Achnanthidium
- A.binodis
- A.delicatula
- A.hauckiana
- A.minutissima
- A.oblongella
- A.plonensis
Amphora
- A.australiensis
- A.coffeaeformis
- A.fontinalis
- A.holsatica
- A.ovalis
- A.obtusa
Aneumastus
Anomoeoneis
Aulacoseira
Bacillaria
Brachysira
- B.serians
- B.styriaca
- B.vitrea
Caloneis
Camphylodiscus
Cocconeis
Craticula
Ctenophora
Cyclotella
- C.atomus
- C.menenghiniana
- C.stelligera
- C.striata
Cymbella
- C.lanceolata
- C.microcephala
- C.similis
- C.tumida
Denticula
Diadesmis
Diploneis
- D.chersonensis
- D.ovalis
- D.smithi
- D.vacillans
Encyonema
Entomoneis
Epithemia
Eunotia
- E.camelus
- E.curvata
- E.eurycephaloides
- E.exigua
- E.flexuosa
- E.formica
- E.incisa
- E.pectinalis
- E.trigibba
Fallacia
Fragilaria
- F.brevistriata
- F. punctata
- F.rumpens
- F.vaucheriae
Frustulia
- F.megaliesmontana
- F.rhomboides
- F.vulgaris
Gomphonema
- G.acuminatum
- G.affine
- G. lanceolatum
- G.parvulum
- G.truncatum
- G.undulatum
Gyrosigma
- G. balticum
- G.kutzingii
- G.spenceri
Hantzschia
|
Hyppodonta
Luticola
Martyana
Mastogloia
- M.braunii
- M.elliptica
- M.smithii
Melosira
M.moniliformis- M.nummuloides
- M.varians
Meridion
Navicula
- N.accomoda
- N.cancellata
- N.cincta
- N.cryptocephala
- N.elginensis
- N.festiva
- N.gastrum
- N.radiosa
- N.ryhnchocephala
- N.salinarum
- N.stigmatifera
- N.symmetrica
- N.tripunctata
- N.virudila
Neidium
Nitzschia
- N. acicularis
- N.amphibia
- N.closterium
- N.dissipata
- N.gracilis
- N.linearis
- N.lorenziana
- N. microcephala
- N.obtusa
- N.palea
- N.paleacea
- N.sigma
Paralia
Pinnularia
- P.subcapitata
- P.borealis
- P.divergens
- P.gibba
- P.maior
- P.microstauron
- P.viridis
Pleurosigma
- P.elongatum
- P.longum
- P.salinarum
Rhopalodia
- R.brebissoni
- R.gibba
- R.gibberula
- R.musculus
Sellaphora
Skeletonema
Stauroneis
- S.anceps
- S.dubitabilis
- S.pachycephala
- S.spicula
Staurosira
- S.construens
- Stenopterobia
- S.sigmatella
Surirella
- S.angusta
- S.ovalis
- S.tenera
Synedra
Tabellaria
Tabularia
Thalassionema
Thalassiosira
Tryblionella
- T.apiculata
- T. circumsuta
- T. hungarica
- T.levidensis
- T.victoriae
|
Meta Data Record
<TITLE>A Guide to Diatoms as Indicators of Urban Stream Health</TITLE>
<META NAME="DC.Title" CONTENT="A Guide to Diatoms as Indicators of Urban Stream Health">
<META NAME="DC.Creator" CONTENT="Jacob John. School of Environmental Biology Curtin University of Technology">
<META NAME="DC.Type" CONTENT="text">
<META NAME="DC.Date" CONTENT="2000">
<META NAME="DC.Format" CONTENT="text/html">
<META NAME="DC.Coverage" CONTENT="Australia">
<META NAME="DC.Description" CONTENT="The ultimate monitor of aquatic systems such as urban streams is aquatic life itself. No other single taxonomic group of aquatic organisms is as sensitive as diatoms to changes both in water quality and habitats. For any biomonitoring program to be successful, the biomonitors need to be identified accurately. Lack of practical guides to identify diatoms is generally acknowledged as a problem limiting their use as biomonitors in Australia. This publication is an attempt to address this problem.">
<META NAME="DC.Relation" CONTENT="This report describes the outcomes of a research project conducted under the Urban Research and Development sub-program of the National River Health Program (NRHP). The NRHP is an on-going national program established in 1993, managed by the Land and Water Resources Research and Development Corporation (LWRRDC) and Environment Australia. Its mission is to improve the management of Australia's rivers and floodplains for their long-term health and ecological sustainability">
<META NAME="DC.Source" CONTENT="Environment Australia community Information Unit">
<META NAME="DC.Subject" CONTENT="Water, rivers">
<META NAME="DC.Publisher" CONTENT="Land and Water Resources Research and Development Corporation. GPO Box 2182 Canberra ACT 2601">
<META NAME="DC.Publisher" CONTENT="Published Electronically on au.riversinfo.org by the Environmental Information Association (Incorporated) with the permission of LWRRDC and Environment Australia.">
<META NAME="DC.Rights" CONTENT="Copyright (©) LWRRDC">
<META NAME="DC.Identifier" CONTENT="LWRRDC Occasional Paper 14/99 (Urban Sub Program, Report No.7) 2000 ISBN 0 642 26764 2">
<META NAME="DC.Identifier" CONTENT="http://au.riversinfo.org/library/nrhp">
<meta name= "DC.Language" content = "en">
 Riversinfo Australia (au.riversinfo.org): Publication Information:
Riversinfo Australia (au.riversinfo.org): Publication Information:
- Webserver publication date (version): Saturday, April 20th 2019.
- The publication of material on au.riversinfo.org was supported by the Commonwealth Government via the National Landcare Program and the National River Health Program.
- End users are advised to expect variations in formatting and layout depending on both the format of the original material, as supplied by the authors, and the end users particular software and hardware configuration(s).
- In the case of variation between this document and a hard copy original the original takes precedence.
- Except when otherwise stated, in writing, information provided by au.riversifno.org is provided "as is" without warranty of any kind to anyone. The entire risk as to use of the information rests with the user. Should the information prove defective, the user of the information assumes the cost of all necessary repair or correction. In no event, unless agreed to in writing, will anyone associated with au.riversinfo.org be liable for any damages arising out of the use or inability to use the information, even if au.riversinfo.org has been advised of the possibility of such damages.
- As of January 2004 au.riversinfo.org is archived under Precision Info.
Copyright ©
- Authors and the public are reminded that the purpose of au.riversinfo.org is to compile and reference Internet information about rivers in an easily accessible and comprehensive manner to assist all those seeking to preserve and publicize Australia's Rivers.
- au.riversinfo.org has no interest in acquiring or assuming exclusive copyright over any authors material published on au.riversinfo.org. The copyright of material appearing on au.riversinfo.org rests with existing copyright holders who give approval for publication on au.riversinfo.org.
- The copyright status of articles is generally stated within the article and those seeking to reproduce the article further should, if required, seek permission from the copyright holder prior to any further publication or circulation.
- Where no explicit copyright statement has been made copyright will be assumed to be held by the author of the article.
- au.riversinfo.org principally converts and marks up existing material from various formats. The au.riversinfo.org content delivery scripts, mark up code and presentation are copyright.
- Consistent with the aims stated in point 1 and provided you do not attempt to substantially reproduce large sections of the au.riversinfo.org Internet site you may, provided you have the permission of the copyright holders, copy and distribute verbatim copies of the files appearing on au.riversinfo.org provided that you conspicuously and appropriately publish on each copy an appropriate notice acknowledging the source of the material.
- Consistent with the aims stated in point 1 and provided you do not attempt to substantially reproduce large sections of the au.riversinfo.org Internet site you may, provided you have the permission of the copyright holders, copy and distribute modified versions of the files appearing on au.riversinfo.org provided that you conspicuously and appropriately publish on each copy an appropriate notice acknowledging the source of the material and stating the date and nature of any changes you may have made to the material.
- If you systematically copy and or re-distribute information (or modified information) originating from au.riversinfo.org then you must:
- Accompany the distribution with a written offer to transfer the information to any third party, wishing to use it in a manner consistent with the aims stated in point 1, a complete machine-readable and interchangeable copy of the corresponding material.
- Any charges to the third party arising as a result of the transfer must not exceed the cost of your physically performing the transfer.
- The offer must be valid for at least three years from the date of your last systematic distribution of the material.
- Acceptable machine readable and interchangeable file formats include .html and or .txt and or .rtf formats. PDF format is NOT considered an acceptable machine interchangeable file format.
- You must cause any work that you distribute or publish, that in whole or in part contains information derived from au.riversinfo.org, to be available under the principles of these same distribution terms
- Third parties receiving material deriving from au.riversinfo.org should be advised that they will be bound by the principles of these distribution terms
Acknowledgments
Current at December 2002
 au.riversinfo.org has been developed to facilitate public access to scientific and technical information on Australia's Rivers. Much of this material was paid for using public funds and is therefore owned by the Australian people. Environment Australia have been instrumental in making available all the publications from:
au.riversinfo.org has been developed to facilitate public access to scientific and technical information on Australia's Rivers. Much of this material was paid for using public funds and is therefore owned by the Australian people. Environment Australia have been instrumental in making available all the publications from:
- The National River Health Program
- The Ausrivas Assessment Scheme
 The au.riversinfo.org Internet site is in-part supported by the Natural Heritage Trust. The Natural Heritage Trust focuses on five key environmental themes - land, vegetation, rivers, coasts and marine, and biodiversity. The programs of the Natural Heritage Trust play a major role in developing sustainable agriculture and natural resource management, as well as protecting our unique biodiversity through improved management and delivery of resources.
The au.riversinfo.org Internet site is in-part supported by the Natural Heritage Trust. The Natural Heritage Trust focuses on five key environmental themes - land, vegetation, rivers, coasts and marine, and biodiversity. The programs of the Natural Heritage Trust play a major role in developing sustainable agriculture and natural resource management, as well as protecting our unique biodiversity through improved management and delivery of resources.
 The au.riversinfo.org Internet site is in-part supported by Sydney Airport who have co-funded a Natural Heritage Trust supported project intended to develop Internet based environmental information integration and management systems. This will assist in developing an overall perspective on the demands and uses for environmental resources
The au.riversinfo.org Internet site is in-part supported by Sydney Airport who have co-funded a Natural Heritage Trust supported project intended to develop Internet based environmental information integration and management systems. This will assist in developing an overall perspective on the demands and uses for environmental resources